- +1
脑声常谈|基底神经节-脑干互作的运动控制神经环路总结
多种运动的执行与学习涉及分布于整个神经系统的神经元网络。脑干和基底神经节是处理运动信息的关键结构。两者均包含功能特化的神经元群体,这些群体基于轴突投射、突触输入和基因表达进行分层,在高分辨率层面揭示了回路解剖结构与功能的对应关系。这两个结构内的神经元群体形成多步骤处理链,专门负责特定运动的执行;然而,这两个结构之间的连接方式与通信机制才刚刚开始被揭示。脑干和基底神经节还嵌入更广泛的网络及系统级环路中。重要的网络组件包括皮层中的广播神经元、小脑输出神经元和中脑多巴胺能神经元。特定动作相关的回路可被增强、否决,与其他回路协同作用或相互竞争或通过可塑性调整以实现适应性行为。运动系统中这种高度特异的回路组织,是支撑行为特异性的核心要素,同时也为行为灵活性提供了充足的结构基础。
引言
行为是一系列在时间上连续发生的运动事件。神经系统需要在特定情境和特定时刻选择并执行恰当的运动,实时追踪并即时调整该运动,并在未来学会更高效地执行它。要深入理解这些过程,关键在于识别产生特定运动所必需的神经环路,并阐明驱动系统“承诺”执行某一运动而非其他可能运动的环路机制,以及学习如何实现这一运动的神经基础。
脑干是一个横跨后脑与中脑的大型中枢,参与多种生理功能,包括运动控制。后脑是脑干最尾端的部分,紧邻脊髓上方,进一步分为脑桥和延髓。后脑向吻侧过渡为中脑,后者包含多个亚区,例如上丘和下丘。为了通过脊髓运动神经元调控肢体运动,脑干恰好位于高级运动中枢与负责运动执行及感觉反馈的脊髓环路之间的连接枢纽。此外,脑干还参与调控其他行为,例如口面部运动(如胡须抖动和舔舐)以及头眼协调运动,这些行为由位于脑干内部的颅神经运动神经元执行。
在运动控制背景下,向脑干多个区域提供输入的一个重要结构是多层结构的基底神经节系统。该系统横跨前脑与中脑区域,已被证明参与运动的学习、选择与执行决策。近期主要在啮齿类动物中开展的研究,正逐步揭示脑干与基底神经节内部处理不同类型运动信息的具体神经环路。利用表达重组酶的基因工程动物结合交叉病毒工具的应用,极大地推动了作者对脑干和基底神经节组织原则与功能机制的理解。这些主要在小鼠中进行的研究揭示出高度精细的细胞层级结构:特定的神经元亚环路专门处理与特定运动形式及其学习相关的信息。这些发现促使作者提出基底神经节与脑干可能如何相互连接以处理运动相关信息并嵌入更广泛的系统级网络之中。
调控运动速度与方向的脑干神经元
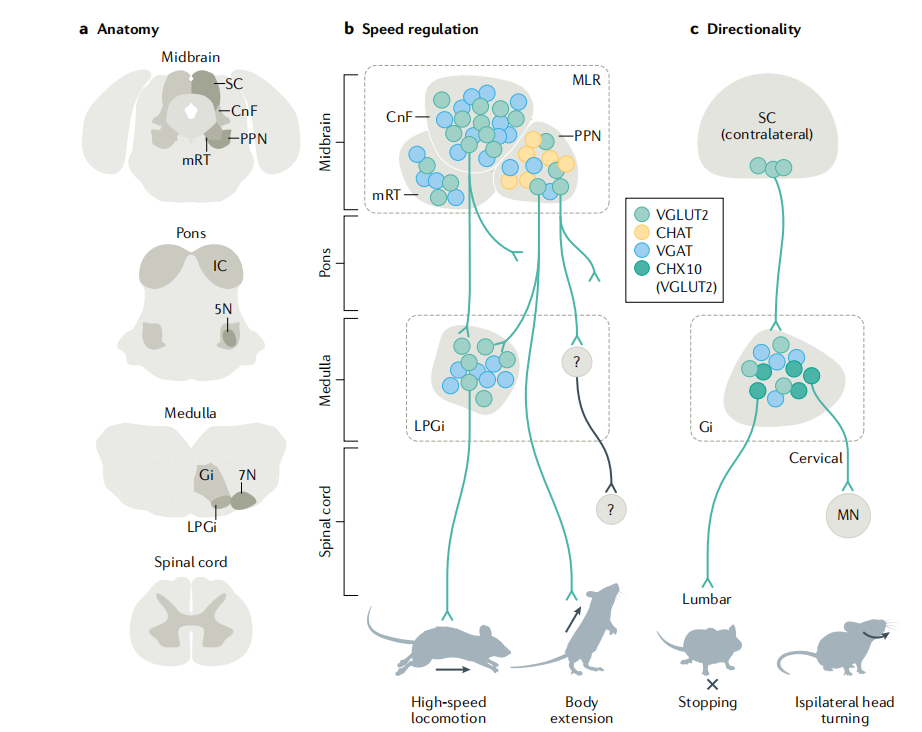
脑干在运动控制中扮演核心角色,尤其在调控肢体参与的两类行为运动的速度与方向性以及不伴随全身位移的前肢精细动作方面。尽管脑干内不同功能的神经元高度混杂,近年研究通过分子标记、投射特异性和功能操控,逐步揭示了其环路组织原则。
在运动速度调控方面,中脑“运动启动区”(MLR)是关键枢纽,包含三个谷氨酸能亚区:脚桥核(PPN)、楔形核(CnF)和中脑网状结构(mRT)。其中,CnF的谷氨酸能神经元对高速运动至关重要,而PPN/mRT群体则更多参与身体伸展(如直立、运动起始)。这些MLR神经元并非全部编码运动状态,部分也参与理毛、抓食等行为,提示其功能多样性。进一步按投射靶点分层可厘清功能:投射至脊髓的PPN/mRT神经元驱动躯体伸展;投射至基底神经节输出核的群体则参与运动选择;投射至延髓的MLR神经元(尤其是CnF→LPGi通路)构成高速运动的核心回路:CnF激活延髓外侧旁巨细胞核(LPGi)的谷氨酸能神经元,后者直接调控脊髓运动程序,实现快速奔跑。相比之下,低速探索性运动的脑干环路尚不明确,但斑马鱼研究提示脊髓可能存在快/慢运动的独立前运动网络。
值得注意的是,单侧激活MLR或LPGi仅引发双侧对称的直线前进运动,无方向改变,除非遇到物理障碍。这表明运动启动信号在脑干或脊髓被对称分配,而环境反馈(如避障)可通过下游环路“否决”正在进行的动作,体现运动执行的可中断性。
在运动方向调控方面,延髓巨细胞核(Gi)中表达转录因子CHX10的谷氨酸能神经元起关键作用。该群体可分为不同投射亚型:颈段投射型:单侧激活引发同侧头部转动,可能通过直接兴奋颈部运动神经元;腰段投射型:激活导致运动减速,但无转向;局部投射型:生理特性不同,功能尚不明。
目前存在两种转向模型:一是不对称停止模型,单侧激活CHX10⁺ Gi神经元抑制同侧运动,对侧继续前行,形成同侧弯曲;二是头-身协调模型,先快速转头,再减速调整身体朝向。上丘深层数神经元(尤其PITX2⁺群体)向对侧Gi提供输入,可能是方向指令的重要来源,而中脑对姿势状态的监测也可能参与调控。
综上,脑干通过功能分离但空间邻近的神经元模块,分别编码运动的速度与方向,并在脊髓整合为完整行为。这种“多通道并行处理”策略,使看似统一的运动行为实则由多个特异信息流协同构建,既保证执行效率,又保留环境交互的灵活性。未来需进一步解析不同CHX10⁺亚群在自然行为中的协同模式以及对侧转向的神经基础。
用于构建前肢动作的脑干神经元
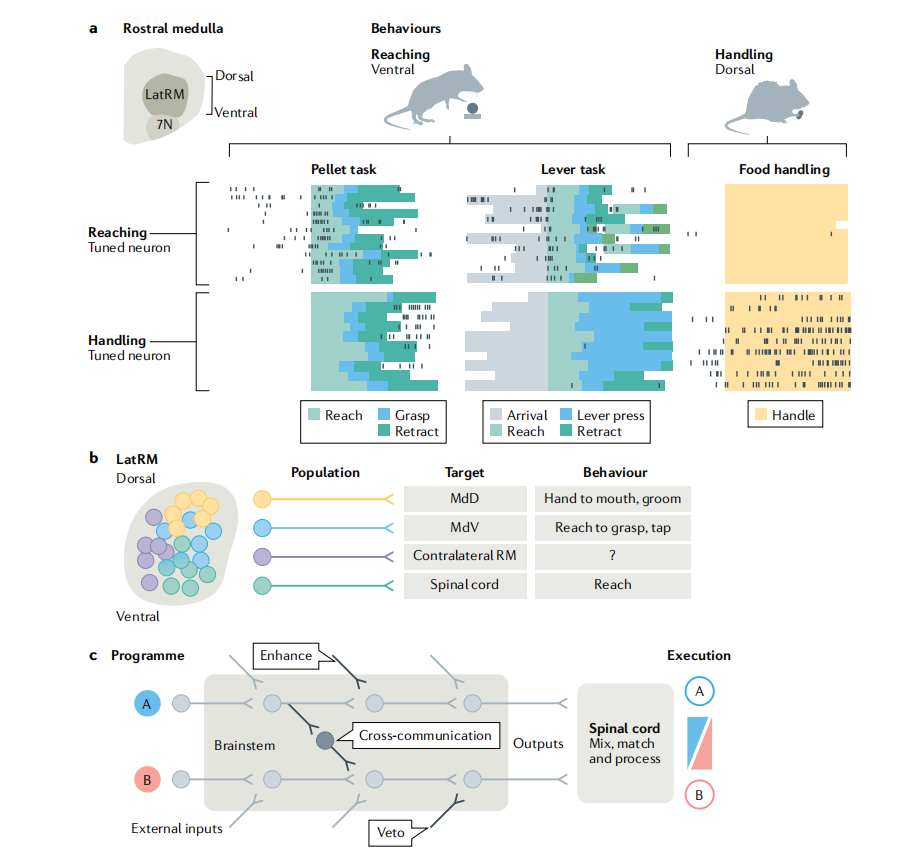
基底神经节与脑干回路的相互作用
脑干中一类专门参与前肢精细动作构建的神经元群体,揭示了其在行为特异性、空间分布和投射功能上的高度组织化。
核心发现:
a. 行为特异性的空间分区
位于延髓头端外侧(latRM)、面神经核(7N)背侧的脑干神经元,根据其参与的前肢行为阶段在空间上分区:
腹侧latRM神经元:特异性编码同侧单侧前肢伸展(如抓取食物颗粒或按压杠杆),但在后续的食物处理(如咀嚼前的手口协调)中不活跃;
背侧latRM神经元:不参与伸展动作,而是在食物抓取完成后的食物处理阶段(如用前肢操纵食物颗粒)被激活。
这些谷氨酸能(兴奋性)latRM神经元对动作的精确性至关重要,抑制它们会导致抓取或操作动作变得笨拙。
b. 投射特异性决定动作类型
通过轴突投射靶点,兴奋性(VGLUT2⁺)latRM神经元可进一步分为至少四类功能亚群:
脊髓投射型:光遗传激活引发单侧前肢伸展;
投射至延髓网状结构背侧部(MdD,最背侧):激活诱发手到嘴动作(hand-to-mouth)和理毛等复杂手指运动;
投射至延髓网状结构腹侧部(MdV):激活引发抓握(reach-to-grasp)和轻敲(tap)动作;
c. 脑干运动环路的通用组织原则
一个概念模型:脑干并非简单传递运动指令,而是对来自上游(如皮层、基底节)的“动作计划”进行动态加工。
假设有两个基本运动程序 A 和 B;
它们在通过脑干时,可被外部输入(如感觉反馈、动机信号)或脑干内部环路(如增强、抑制、“否决”或跨通道交互)所调制;
最终输出到脊髓的信号是经过整合的混合版本;
脊髓再将这些下行信号“混合搭配”生成最终行为,可以是纯A、纯B,或A与B的任意组合。
这一策略使神经系统既能执行标准化动作模块,又能根据情境灵活组合,实现丰富多样的前肢行为。
脑干latRM区域通过空间分区(背-腹轴)和投射特异性(脊髓 vs. 延髓不同亚区),将前肢运动分解为多个可独立调控的功能单元。这种“模块化+可重组”的架构使脑干不仅能执行特定动作(如伸展、抓握、手口协调),还能支持行为的精确性与灵活性,是理解精细运动控制的关键环节。
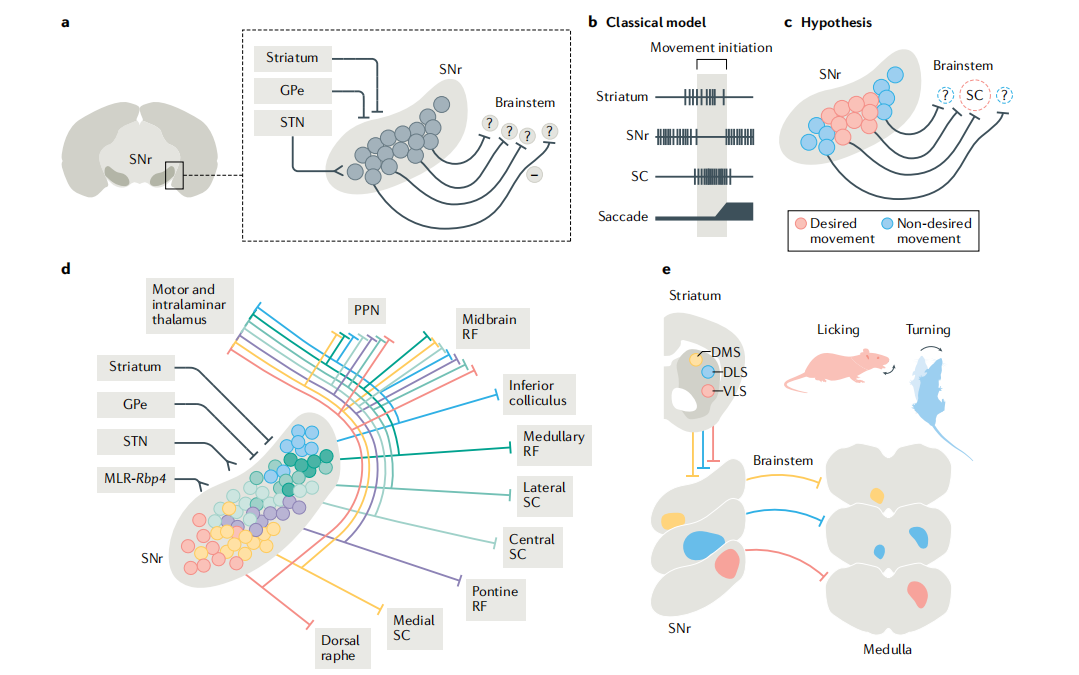
基底神经节(以黑质网状部SNr为核心)与脑干的回路连接及功能分工:
基底神经节通过其核心输出核团黑质网状部(SNr)与脑干形成高度特异的功能回路,实现对运动的精准调控。SNr接收来自纹状体、苍白球外侧部(GPe)和丘脑底核(STN)的输入并通过多条GABA能抑制性投射通路靶向脑干多个区域(如上丘、脚桥核PPN、中脑网状结构等),部分通路功能尚待明确。
传统“去抑制”模型认为,运动启动依赖于层级抑制的解除:纹状体激活后抑制SNr,从而解除SNr对脑干上丘(SC)的抑制,使上丘得以激活并触发扫视运动。这一机制强调基底神经节通过“释放”预先存在的脑干运动程序来启动行为。
然而,新证据支持更精细的“选择-抑制”模型:SNr内部包含功能分化的神经元亚群:一部分调控目标动作,另一部分抑制非目标动作。通过同时激活目标通路并抑制竞争性通路,SNr实现动作的选择性执行,而非简单地全局去抑制。
这种功能特异性源于SNr投射的高度分区性:不同亚群精准投射至脑干特定核团及丘脑,形成一一对应的输出通道。例如,纹状体背内侧区(DMS)、背外侧区(DLS)和腹外侧区(VLS)分别通过各自的SNr亚群,选择性调控舔舐、躯体转向等特定动作,构成“纹状体分区→SNr亚群→脑干功能区”的闭环回路。
综上,基底神经节不仅通过经典的“去抑制”机制启动运动,更借助SNr神经元亚群的投射特异性与功能分化,在脑干层面实现动作的精准选择、抑制与分区调控,从而支撑复杂、灵活的行为输出。
纹状体回路中的动作特异性
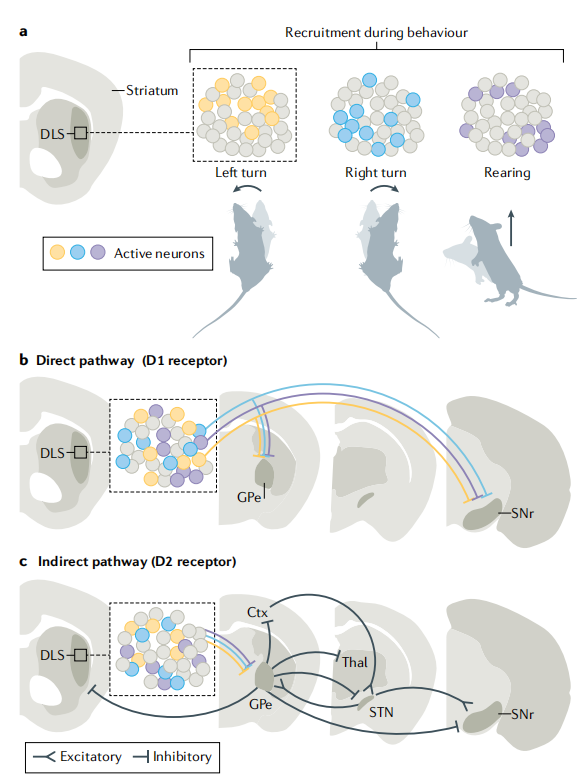
纹状体背外侧区(DLS)的神经元亚群如何通过直接/间接通路实现动作特异性调控:
a 行为中的神经元募集(动作特异性激活)
纹状体背外侧区(DLS)包含不同的神经元亚群,在不同行为中特异性激活:
原标题:《脑声常谈|基底神经节-脑干互作的运动控制神经环路总结》
本文为澎湃号作者或机构在澎湃新闻上传并发布,仅代表该作者或机构观点,不代表澎湃新闻的观点或立场,澎湃新闻仅提供信息发布平台。申请澎湃号请用电脑访问http://renzheng.thepaper.cn。



- 报料热线: 021-962866
- 报料邮箱: news@thepaper.cn
互联网新闻信息服务许可证:31120170006
增值电信业务经营许可证:沪B2-2017116
© 2014-2026 上海东方报业有限公司

