- +1
纽结的形态——“结”论的历史、应用与未解
纽结遍布我们周遭的世界,既可以是缠绕的线缆、鞋带,也能是脱氧核糖核酸(DNA)的复杂构造。纽结绝非单纯的麻烦之物,它蕴藏深邃又优美的数学性质,与几何学、拓扑学、物理学、生物学都有着千丝万缕的关联。

本次讲座,Alain Goriely教授将带大家走进奇妙的纽结理论(knot theory)世界,讲解对纽结的数学研究如何帮助我们理解、预判其形成、稳定性与形态变化规律。
作者:Alain Goriely(阿兰·戈里利教授)2026-6-9
译者:zzllrr小乐(数学科普公众号)2026-6-23
主持人开场
各位晚上好,欢迎来到巴恩斯礼堂,欢迎莅临格雷沙姆学院(Gresham College)。我是克里斯汀・古德费洛(Christine Goodfellow),本院学术项目主管。今晚我十分荣幸为大家带来本场讲座:纽结的形态 —— 从脱氧核糖核酸(DNA)到鞋带、太阳耀斑(solar flare)。本场是《自然几何与万物形态》系列讲座第六场,也是收官之作。
接下来,请掌声有请几何学教授阿兰・戈里列(Alain Goriely)。
阿兰教授讲座内容全文

非常感谢克里斯汀,也感谢所有到场、线上收看的观众。今天的内容十分特别。每年收官讲座我都会挑选一个兼具深度数学内涵、应用场景丰富的主题。大约25年前我攻读博士后时便开始研究纽结(knot,也简称结)相关课题,此后多年持续探索它各维度特性,今天和大家分享我的研究成果。
随处可见的纽结

说起结,生活中随处可见。低头看看鞋带,或是系领带的人都能见到。但纽结的历史远比人类文字记载更久远,早在文字诞生之前,人类就已掌握打结技艺。航海、外科手术、登山等领域,纽结都是不可或缺的基础工具。

阿什利(Clifford Warren Ashley,1881 - 1947,美国)1944年出版的著作完整梳理了人类打结文明,他耗时11年整理出近4000种绳结。书的序言里他写道:简单的打结动作,是一场无限维度的空间探险。接下来,我将带领大家开启这场探索之旅。

我们身边既有独立绳结,也有乱线缠结(tangle),还有编发这类多股线材交织结构。自然界同样遍布纽结:太平洋鳗鲡(Pacific hagfish)遇天敌时会将自身身体打成结,也依靠结状结构清理体表黏液。
本次讲座会多角度讲解纽结应用。微观层面,结理论是分子生物学、脱氧核糖核酸(DNA)研究的核心工具,至今仍是解析生物发育的关键;微观尺度约10的负8次方米,而宏观宇宙,比如太阳内同样存在纽结结构,太阳耀斑便是典型代表。

太阳爆发耀斑时会释放巨量能量,放大观测可见大量缠绕磁通量管(magnetic flux rope),这类管状结构汇聚磁场,绕公共轴线盘旋,整体尺寸远超地球。

纽结科学的起源便和这类宇宙大尺度缠绕结构息息相关,源头可追溯至1858年赫尔曼・冯・亥姆霍兹(Hermann von Helmholtz)的研究。

我很喜欢展示科学家年轻时的照片,彼时他们满怀求知热忱,而非暮年授课的老者形象。亥姆霍兹的核心发现是涡环(vortex ring),也就是烟圈这类流体涡旋结构。涡线(vortex line)标记流体旋转区域,他证明理想流体中涡旋形态随时间守恒,这也是烟圈形态稳定的原因。
20年前我购置高速摄像机,完成一组实验:两台烟炮喷出两道烟环,烟环能够相互穿插、重新联结。这套流体穿插、重组、相互作用的理论,启发了另一位青年学者 —— 彼时尚未获开尔文勋爵(Lord Kelvin)头衔的威廉・汤姆森(William Thompson)。1860年代末,汤姆森拓展亥姆霍兹的理论,提出以太(ether)涡环原子假说:不同元素对应复杂程度各异的涡环结。该猜想一度引发学界热烈讨论,他的友人、爱丁堡大学教授彼得・泰特(Peter Tait)投入相关研究。

泰特认为,想要区分各类元素,就必须系统分类所有可能的纽结构型,由此开启纽结理论的系统性研究,也是数学结论(mathematical knot theory)的奠基工作。他在著作序言写道:结论或将成为数学重要分支。接下来我们会看到,它不仅是重要分支,更是拓扑学(topology)、曲面几何、形态研究的基础核心,作用等同于数字在各类数学分支中的地位。
下面讲解数学层面结的定义。对数学家而言,纽结的规范定义:三维欧几里得空间内圆周(S¹)的嵌入(embedding),在环境同痕(ambient isotopy)意义下等价。
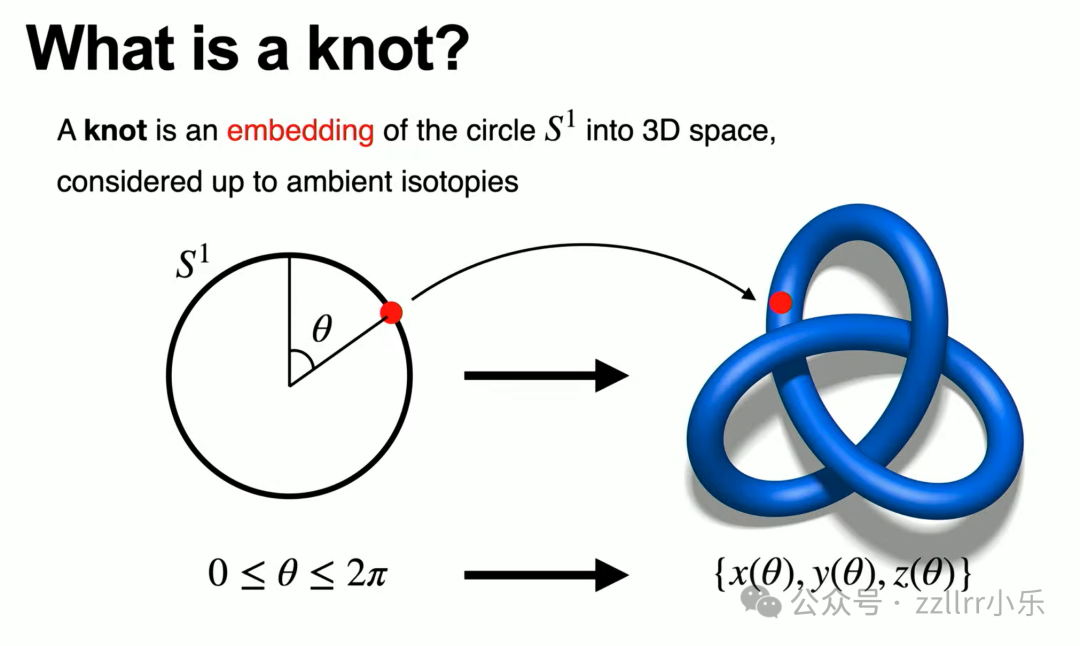
这句话表述晦涩,我们逐层拆解。圆周 S¹ 可通过角度 θ(0 至 2π)参数化,将圆周上每一点映射至三维空间坐标,得到三组连续函数 x、y、z,共同构成一条空间曲线。基础条件:曲线无自交点,线上任意两点不会重合,这便是圆周嵌入,也就是闭合空间曲线,是结的基础形态。
何为环境同痕?简单说,只要不剪断线材、不让线段互相穿过,任意拉伸、扭转得到的构型都属于同一个结。无论简单扭转、小幅拉扯,只要无穿线、无剪断,等价为同一种结,所有这类形态归为同一个等价类(equivalent class)。
举个直观类比:数字1可由一颗橙子、一个苹果代表,实物形态不同,但数值本质一致。同理,不同空间绘制的结图样,只要能无穿线变形互相转化,就是同一个数学结。
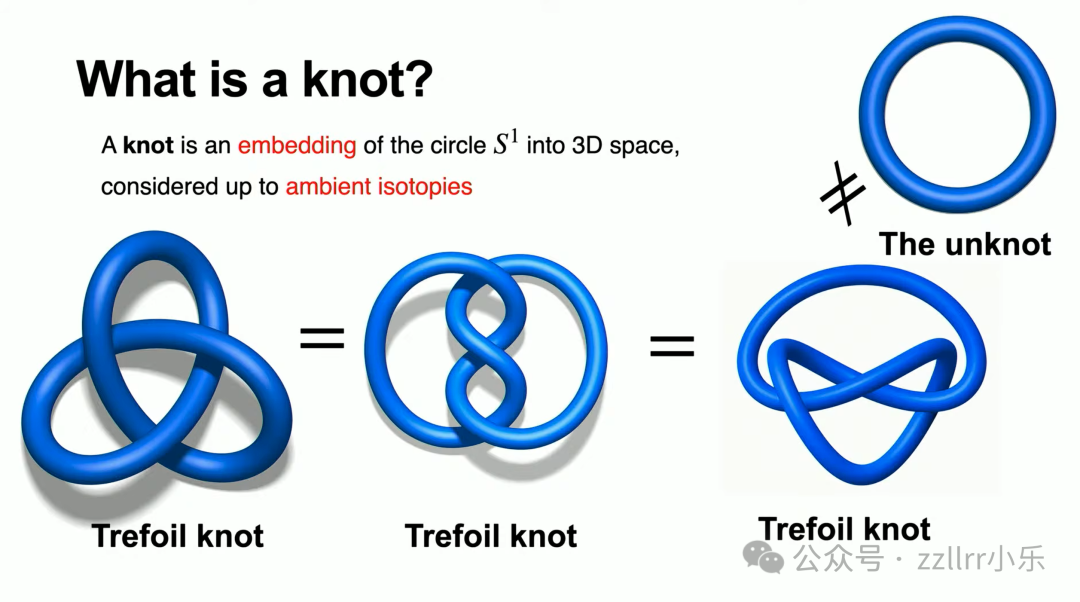
单圈圆环称作无结(unknot),是最简纽结;三叶结(trefoil knot)无法不穿线转化为圆环,二者分属不同等价类。

再举一例,图右侧是无结,左侧看似复杂缠绕曲线,可通过多步无交叉变形完全舒展成圆环,二者等价。
注意:日常活结(overhand knot)不算数学结。
数学定义要求纽结必须首尾闭合。活结比结更通用,因为你有很多方法将
它闭合,因此它是纽结的一种可能推广形式。
如同自然数会拓展至有理数、实数、复数,结理论也有多重拓展方向。
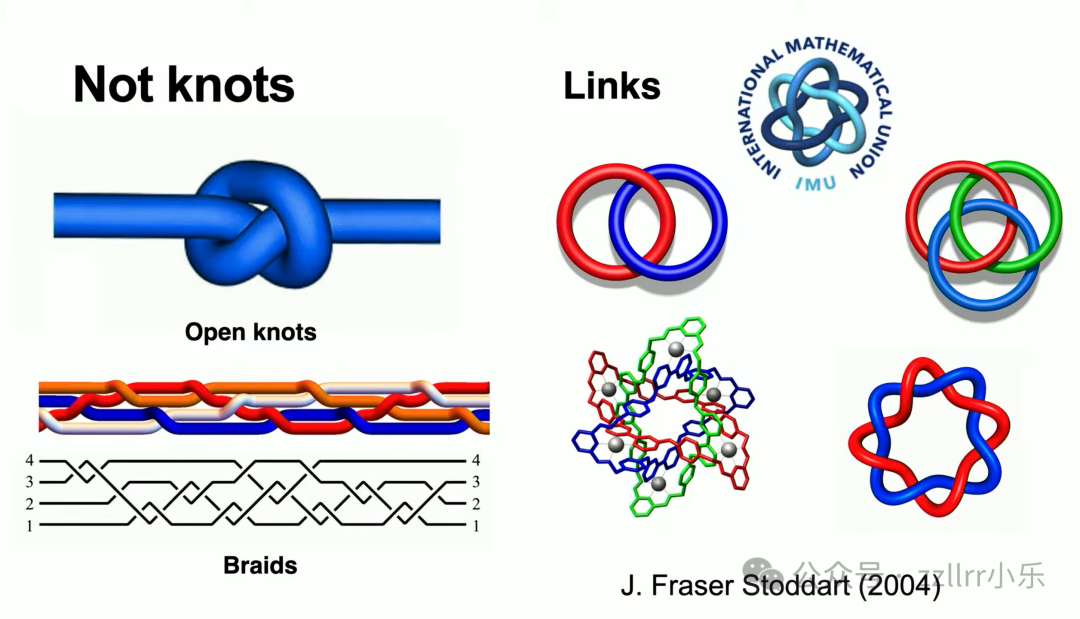
首先是辫(braid):多股线材上下交错编织,编发就是典型案例,纽结是辫的特殊闭合形式;
其次是链环(link),由多条闭合结相互扣合而成。例如 7/9 环面结(Torus knot)可与无结扣合,形成双环链环。链环理论可用于解析蛋白质(protein)分子交联结构。
我们以闭合数学结为核心展开研究。三维曲面难以直接演算,数学家需要可印刷、便于分步推导的二维图示表达,核心方法是结投影图(knot diagram):
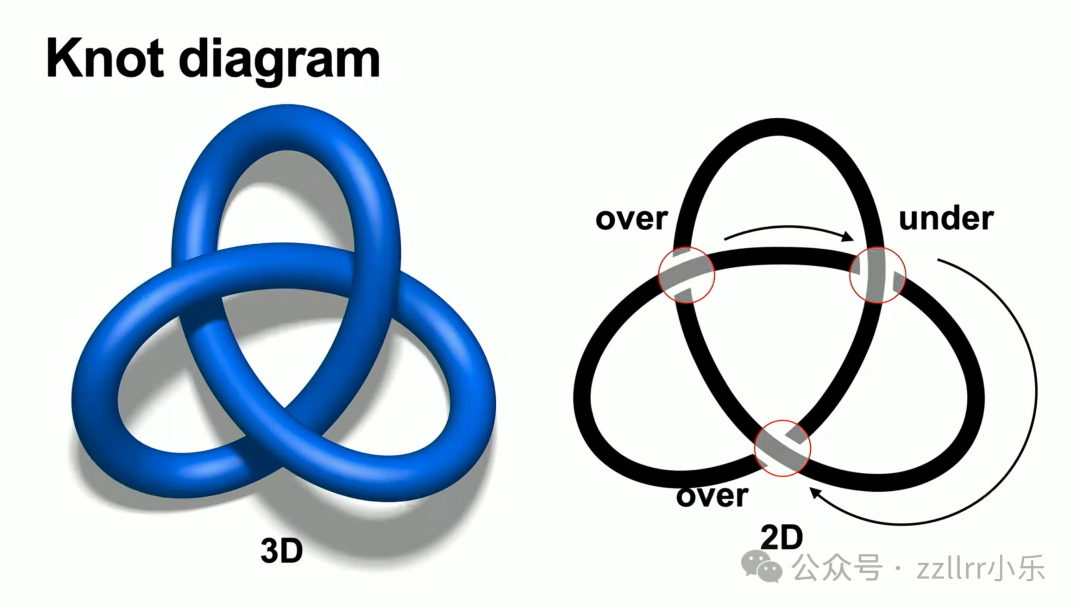
将三维结投影至平面,线段交叉处留白区分上下层,上层线段完整、下层断开,直观区分穿插关系。
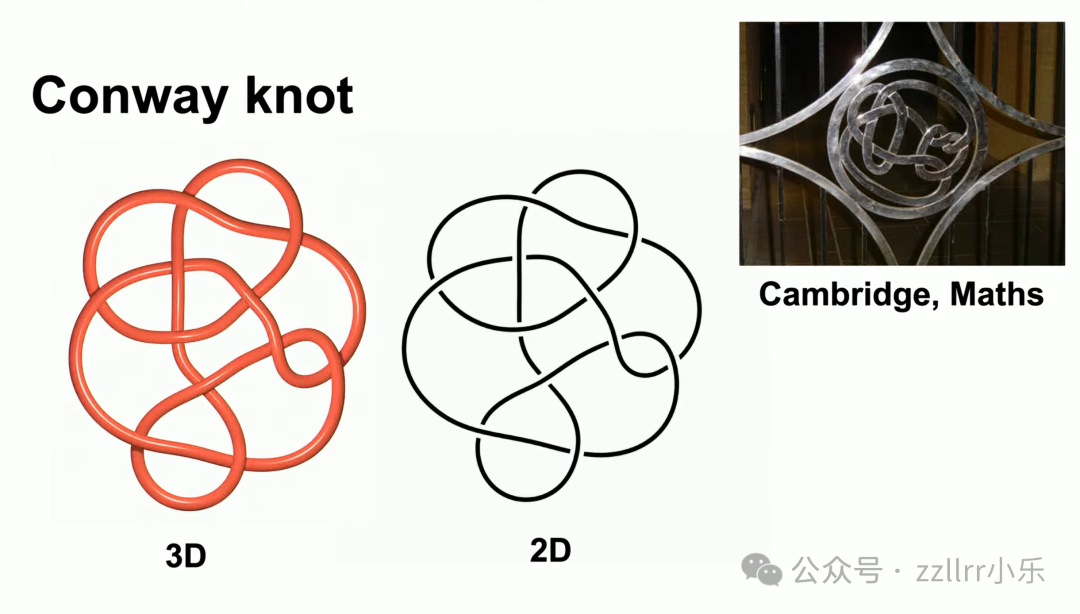
举康威结(Conway knot)示例,图示含 11 个交叉点,平面投影通过留白营造三维视觉效果,纯平面线条也可完整记录全部交叉位置。康威结是知名经典结,剑桥大学数学系、应用数学系与牛顿研究所校区大门印有该纹样。
掌握结图示后,下一步是分类。
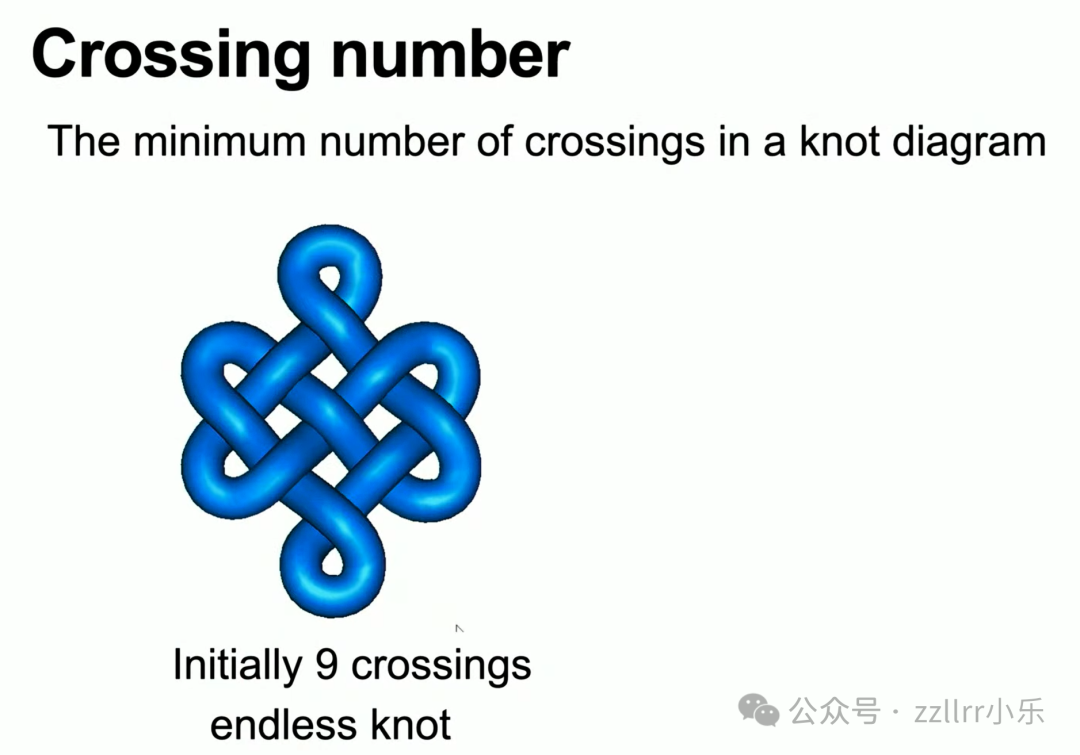
区分结最核心数值是交叉数(crossing number):结所有等效投影中,最少的交叉数量。
无结交叉数为 0;单交叉、双交叉的缠绕曲线均可无穿线舒展为圆环,不存在 1、2 交叉的独立结。
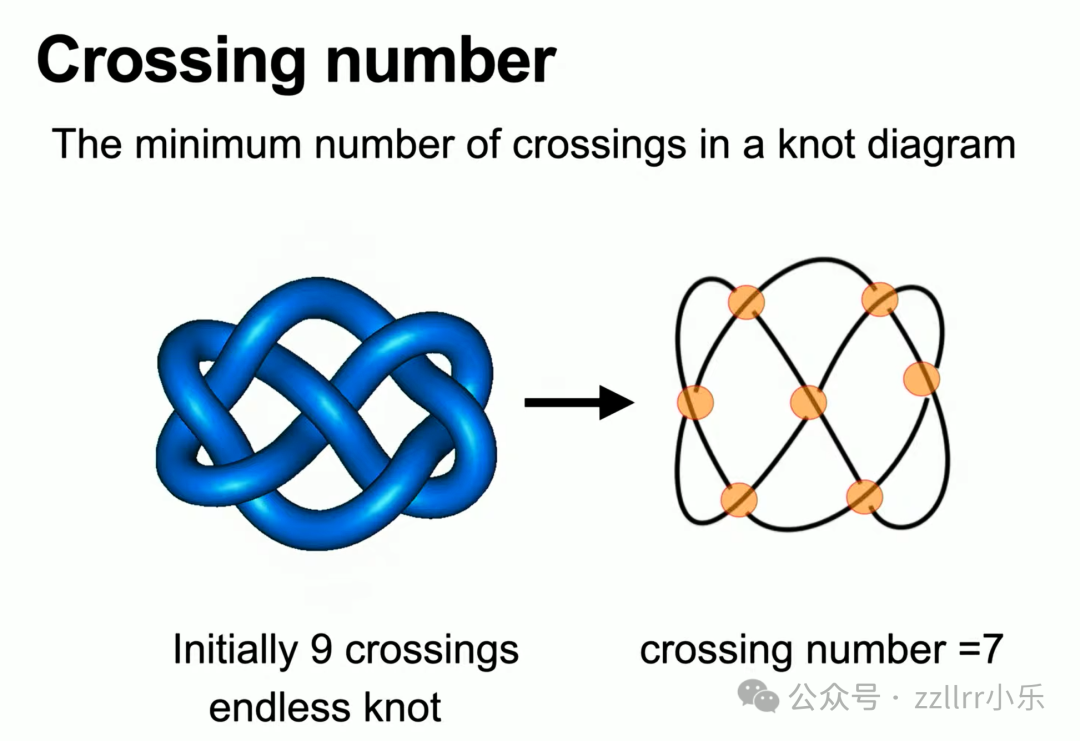
初始九交叉无尽结(endless knot,无尽结,盘长结,吉祥结)看似复杂,通过形变可缩减至 7 个交叉,7 才是它的最小交叉数(该结也称为7₄结,7₄ Knot)。
学界按照最小交叉数依次分类:三交叉、四交叉、五交叉逐一梳理。
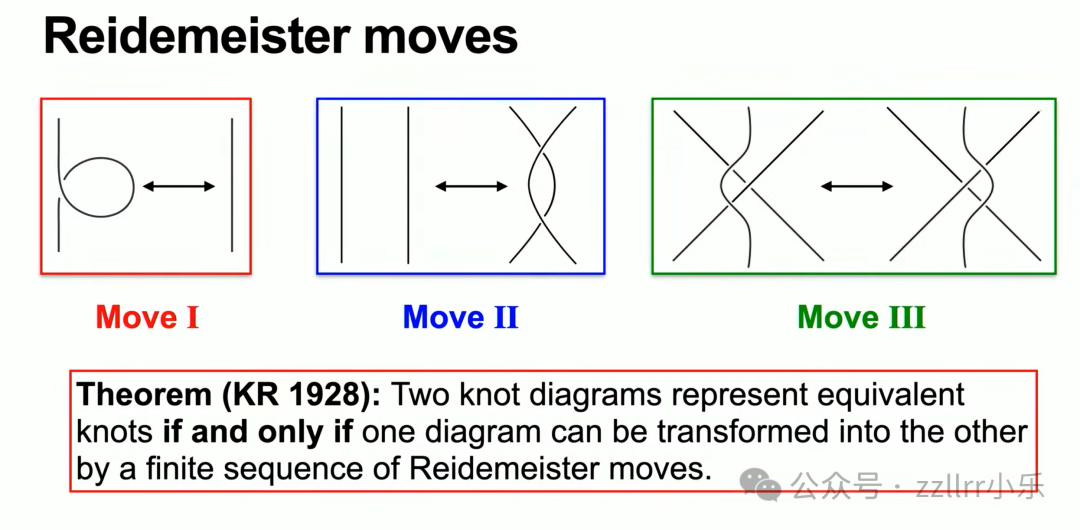
区分两幅投影是否代表同一结,核心工具是瑞德迈斯特变换(Reidemeister move),共三类基础操作:
I类变换(RI):增减单圈小环;
II类变换(RII):两条线段交错,不改变结本质,仅变更图示;
III类变换(RIII):三条线段交错,顶层线段越过另外两条。
瑞德迈斯特定理(Reidemeister theorem):两幅结图示,若可通过有限次三类变换互相转化,则二者为同一结。
一个示例证明:看似复杂的缠绕图形,通过多次一类变换舒展线圈,即可化为无结。
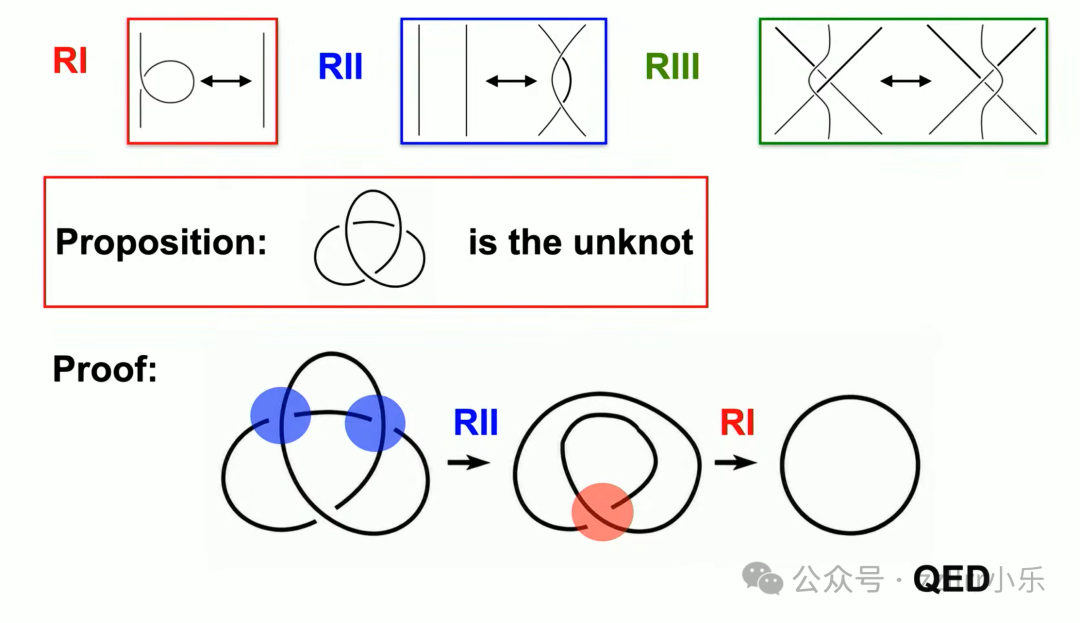
这套证明无数字、无微积分,完全依靠图形变换,是一套可视化严谨数学证明。
再举例“8”字结(figure-eight knot,在系统分类中也记为4₁结):8字结与其镜像能否通过变换等价?
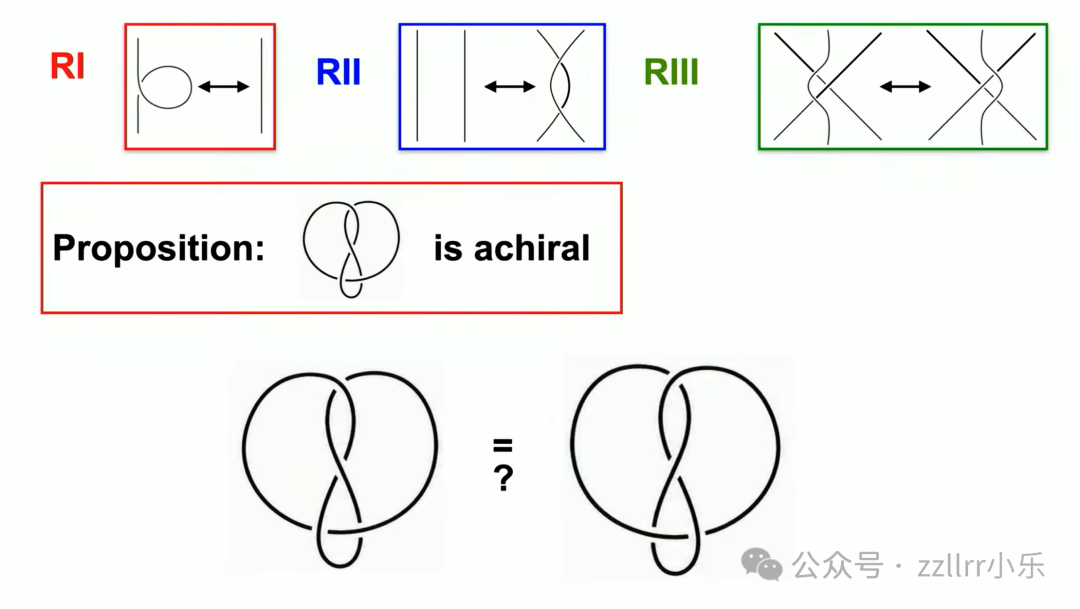
多次二、三类变换重组图示,可完成转化,证明8字结无手性(achiral),镜像为同一结。
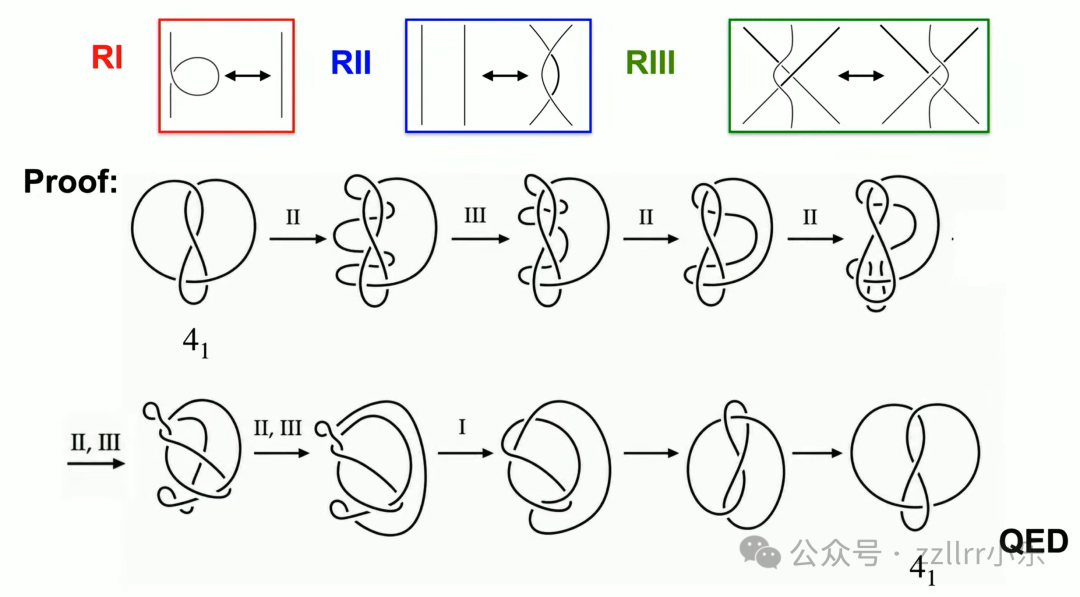
整套证明仅调整平面线条,每一步变换均符合瑞德迈斯特规则,逻辑严密。
纽结的算术运算

如同数字定义加减乘运算,结也可定义复合运算(knot composition),类比整数乘法:
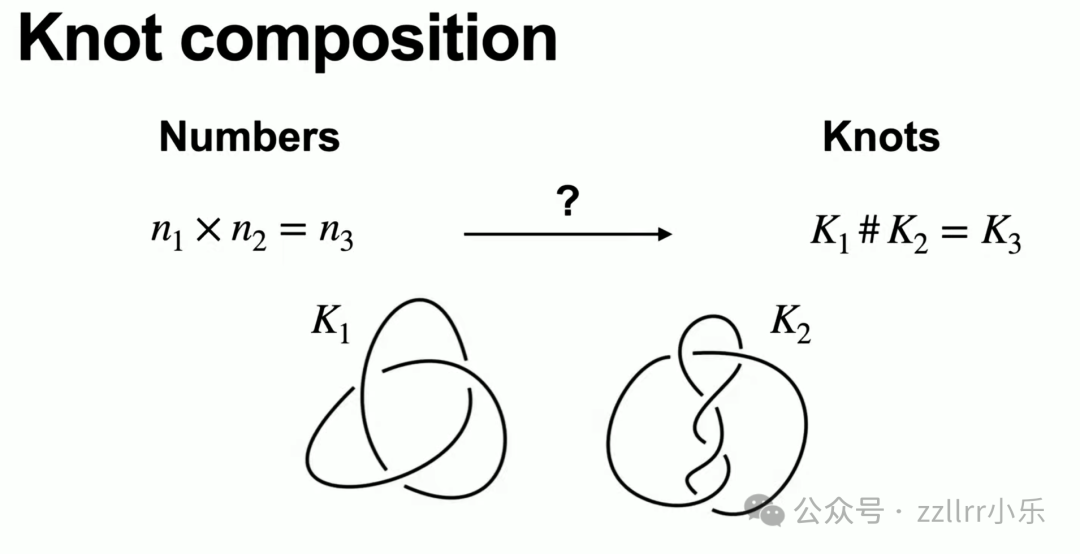
取三交叉、五交叉两种结,各剪断一小段后首尾拼接重组,完成复合。
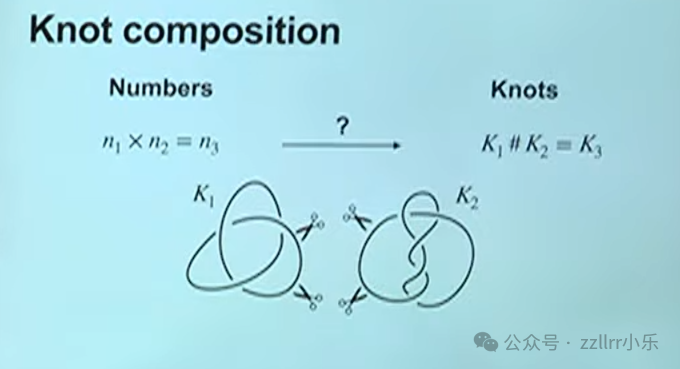
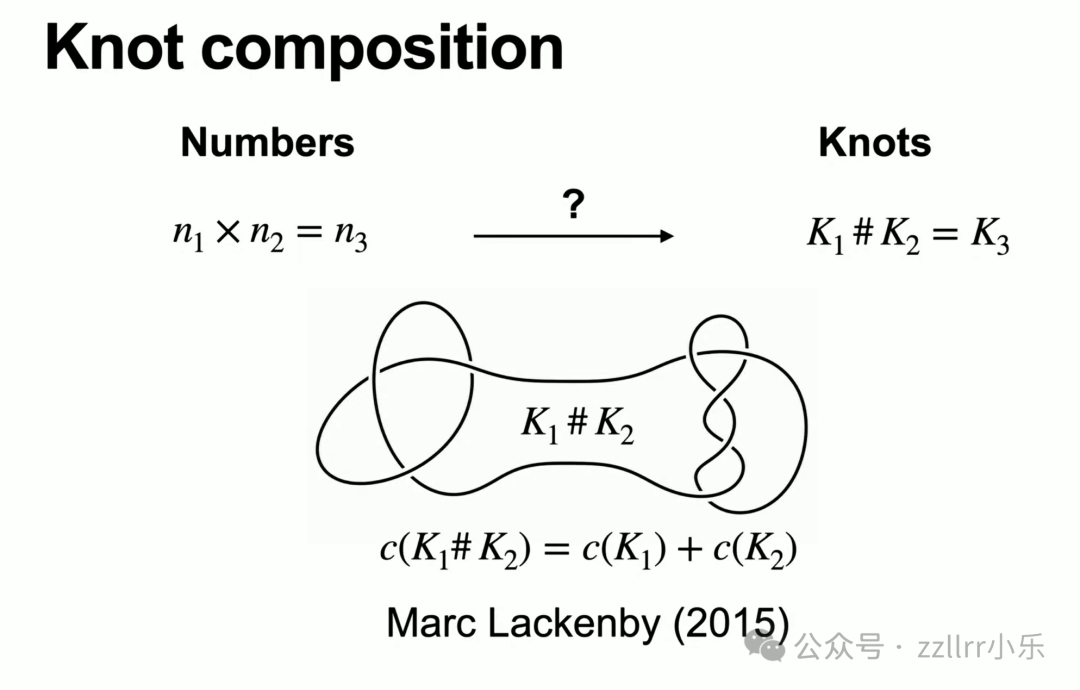
复合后纽结最小交叉数等于原二者交叉数之和,该定理由马克・拉肯比(Marc Lackenby)在十年前证明,他是本院同事、顶尖结论专家。
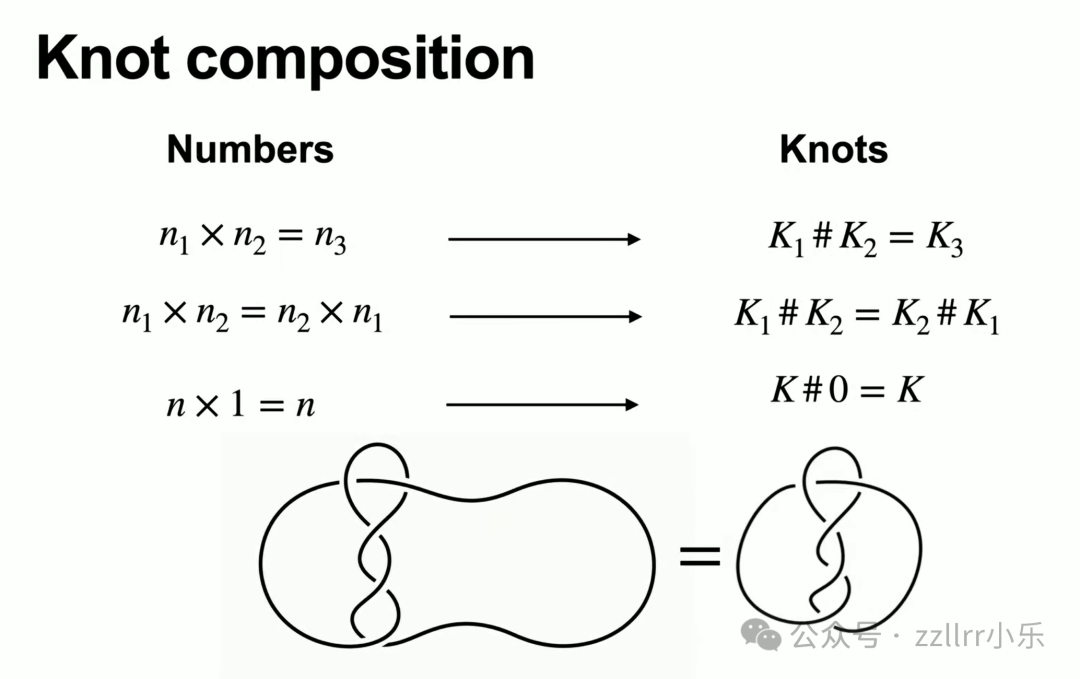
复合运算满足交换律(K₁ 复合 K₂ 等价于 K₂ 复合 K₁);无结是复合运算的单位元,任意结与无结拼接后形态不变。
类比数论素数概念:无法拆分为多个更简单结复合的构型,称为素结(prime knot)。

素结是结的 “基本原子”,所有复杂结均可由素结复合而成。

舒伯特(Horst Schubert)1949年提出纽结的算术基本定理:任意结都能唯一分解为若干素结的复合,类比欧几里得算术基本定理。
素结分类

素结分类统计:
0 交叉:仅无结;
1、2 交叉:无独立素结;
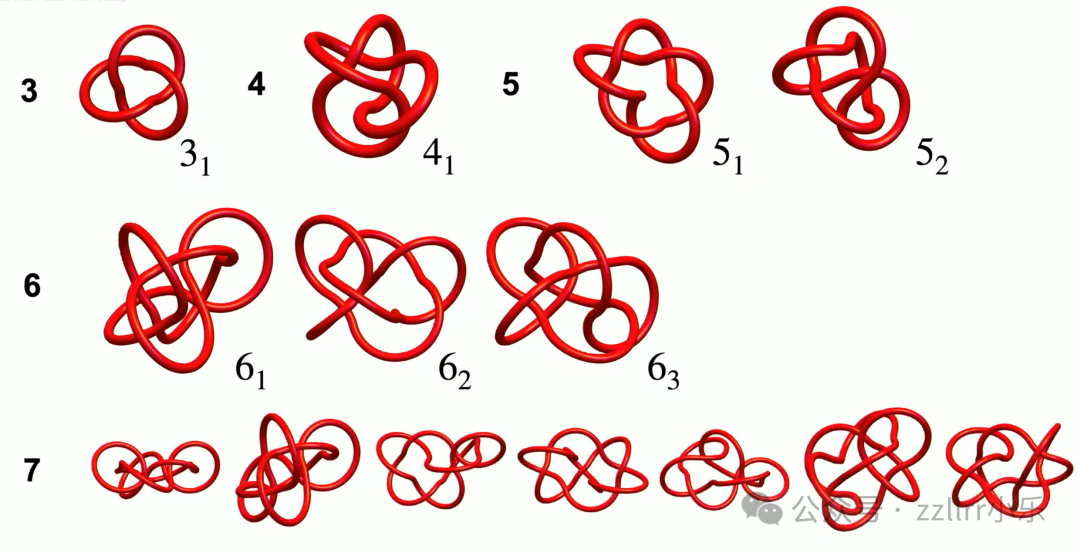
3 交叉:仅三叶结(三叶结镜像无法通过变换等价,1912年才完成严格证明);
4 交叉:仅8字结;
5 交叉:2 种;
6 交叉:3 种;
7 交叉:7 种;
8 交叉:21 种;

9 交叉:49 种;


10 交叉:165 种。



10 交叉编号 10₁₆₁ (上图最后一行第7个)与 10₁₆₂ 曾被泰特(Tait)、利特尔(Little)列入两套独立素结表。

百年后美国律师肯尼斯・珀科(Kenneth Perko)利用业余数学研究证明二者等价,后世称其为珀科对(Perko pair)。
现有免费绘图软件KnotPlot ( https://knotplot.com ),可模拟各类结、支撑学术演算。1900年泰特、利特尔仅统计最高10交叉共 165 种素结;
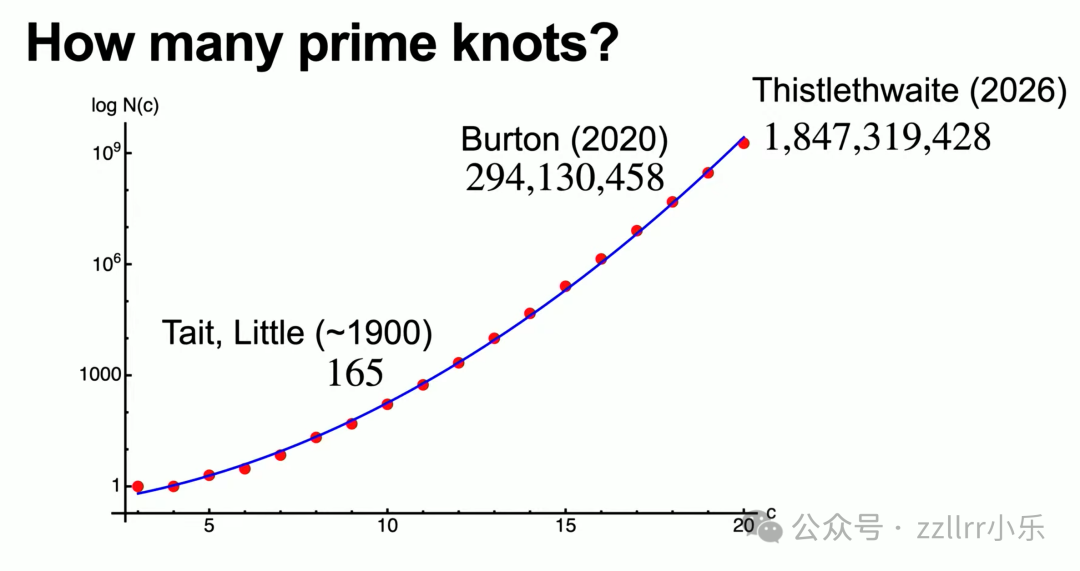
2020年伯顿(Burton)算出19 交叉素结超百万种;
今年莫文·西斯特莱特维特(Morwen Thistlethwaite)刚过完 81 岁生日,证明 20 交叉素结超十亿,21 交叉预估达 210 亿,素结数量随交叉数呈指数级暴涨。
解结

核心开放难题:解结问题。
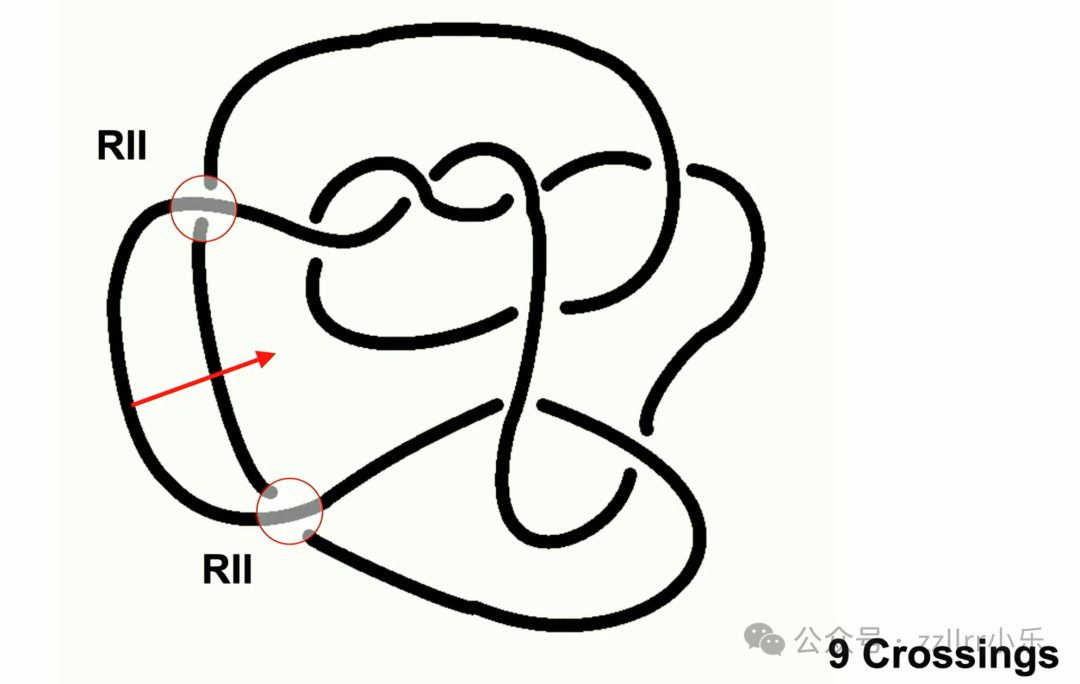
给定结,最少需要多少次瑞德迈斯特变换才能化为无结?
难点在于化简过程中交叉数可能先增多,再减少。

哈肯斯·戈尔迪安结(Hakens Gordian knot)是经典案例,图示 141 个交叉,但本质等价无结。

马克・拉肯比(Marc Lackenby)2015年给出重要理论上界:所需变换次数上限与交叉数的11次幂成正比。
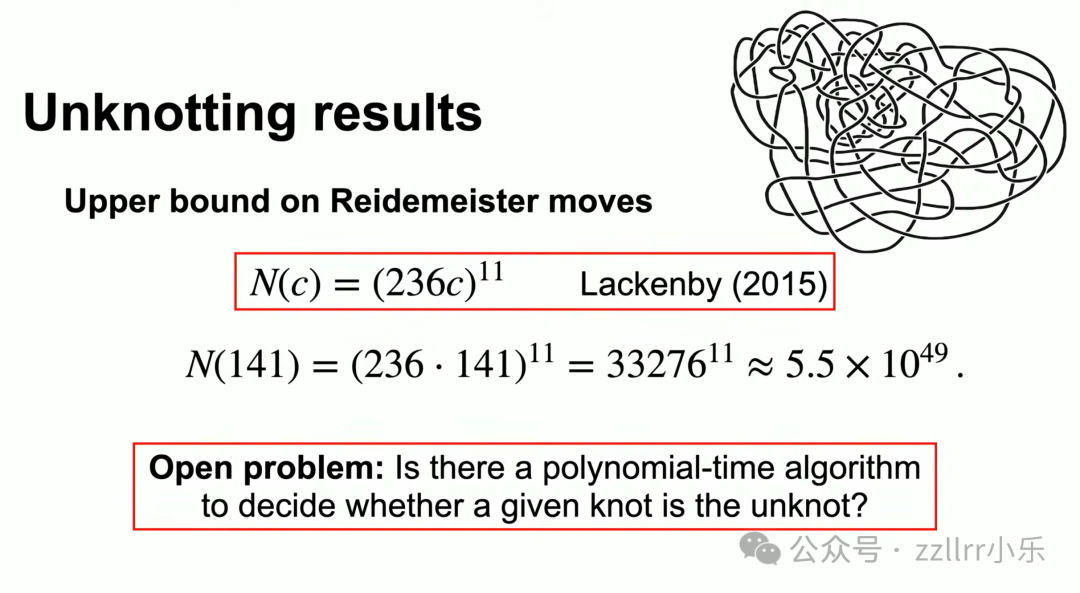
数值极大,若每次变换耗时一纳秒,完成化简所需时间远超宇宙年龄。
该问题属于NP非确定性多项式复杂度问题,学界核心悬题:是否存在多项式时间算法判定结是否为无结?
另一核心操作:交叉变换(crossing change),剪断一段线材、从另一侧穿过再缝合,直接改变结类型。
解结数(unknotting number):将结转化为无结所需最少交叉变换次数。
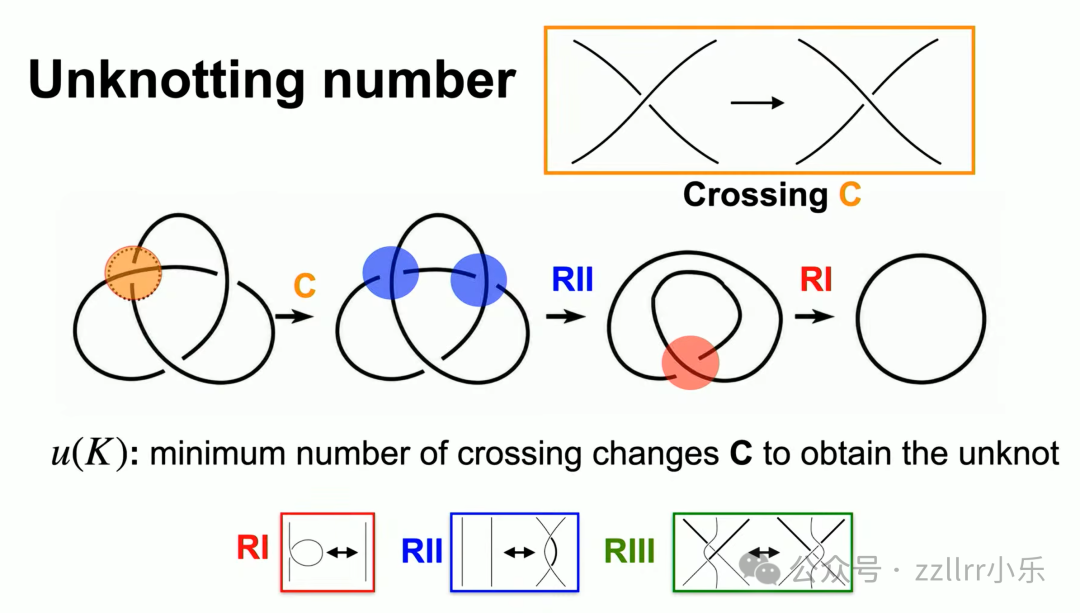
例如五交叉结仅需两次交叉变换即可化为三叶结,三叶解结数为 1。该数值极难精确求解,学界有猜想:非平凡交叉变换必然改变结等价类,但至今无严谨证明。
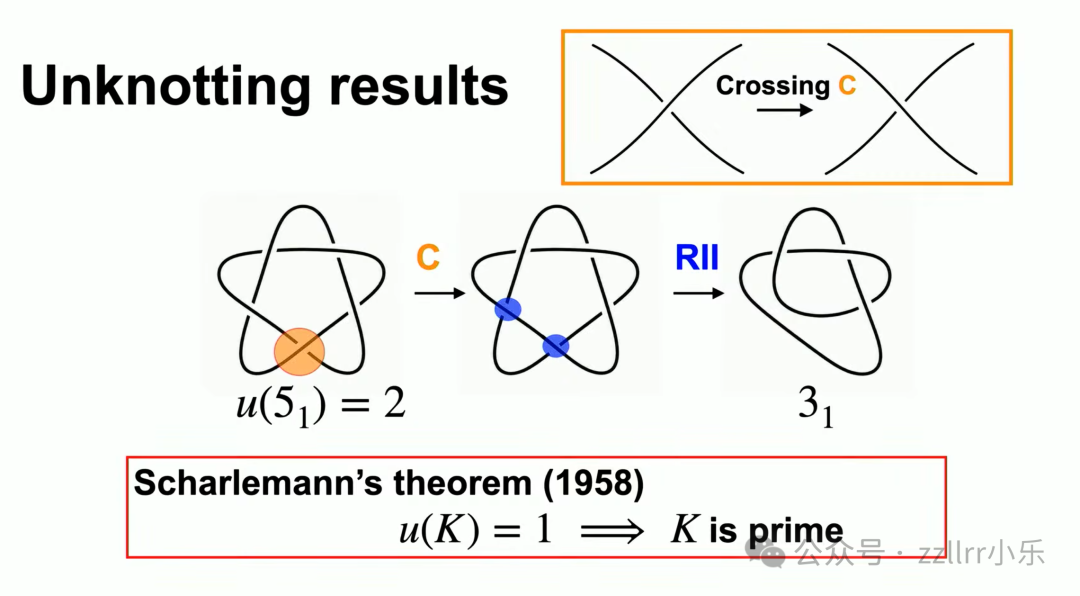
沙勒曼定理(Scharlemann's theorem,1958):若单次交叉变换可将结化为无结,则该结必为素结;反之不成立,五交叉结可佐证。

传说亚历山大大帝斩断戈尔迪乌姆结,或许恰好符合该定理条件,一刀完成交叉变换,再通过瑞德迈斯特变换彻底解结。
结现实应用:脱氧核糖核酸(DNA)

上世纪八九十年代,学界掀起纽结理论分子生物学应用热潮。
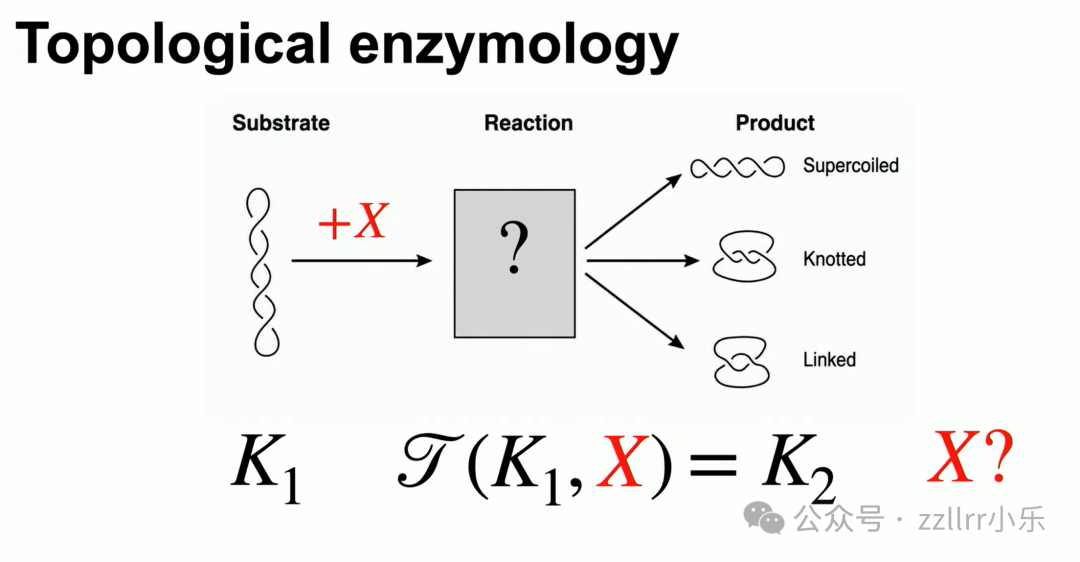
1971年王倬(James C. Wang,1936 -)取得突破性发现:脱氧核糖核酸(DNA)是双螺旋(double helix)结构,携带生物遗传信息,两条互补链。细胞分裂时双螺旋必须完全拆分,才能复制遗传物质。单纯拉扯双链只会加剧缠绕,细胞无巨型外力拆解超长DNA(单条长达2米),生物依靠 拓扑异构酶(topoisomerase)完成解缠:
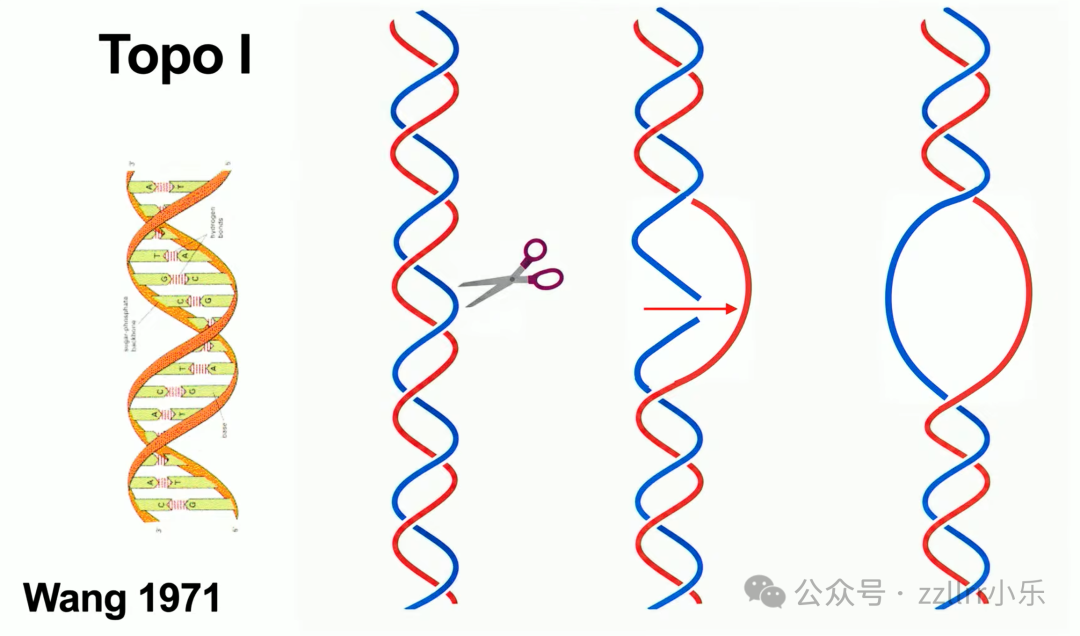
一型拓扑异构酶(Topo I):切断单链,穿过缠绕区域后重接,每次减少一组螺旋扭转。
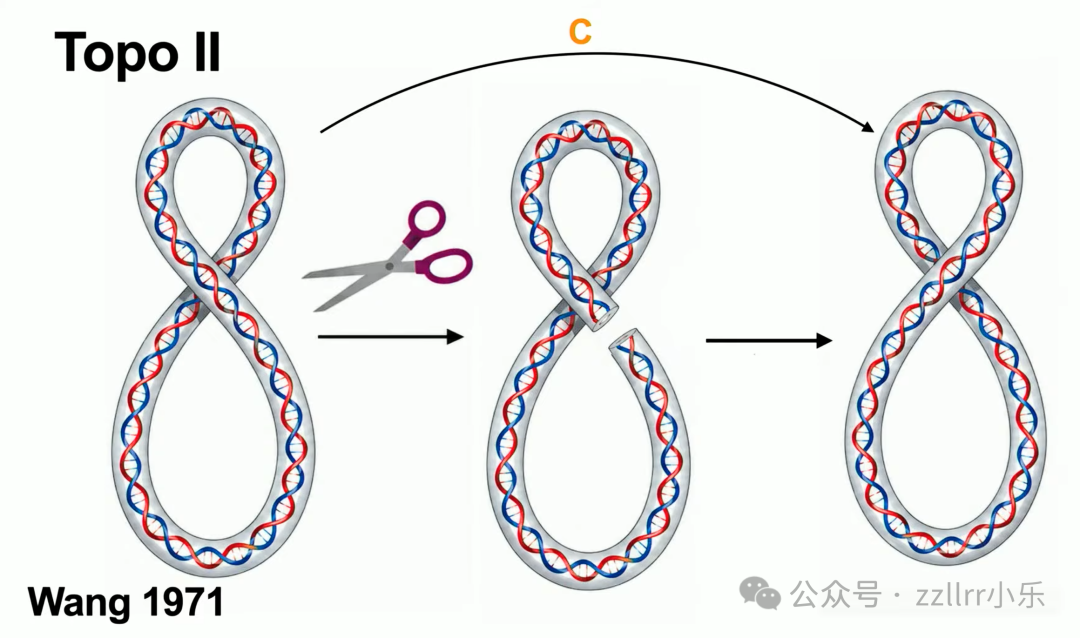
二型拓扑异构酶(Topo II):同时切断双链,整条DNA穿过缝隙后缝合,可拆分互锁环形 DNA(catenane,索烃,链环)。
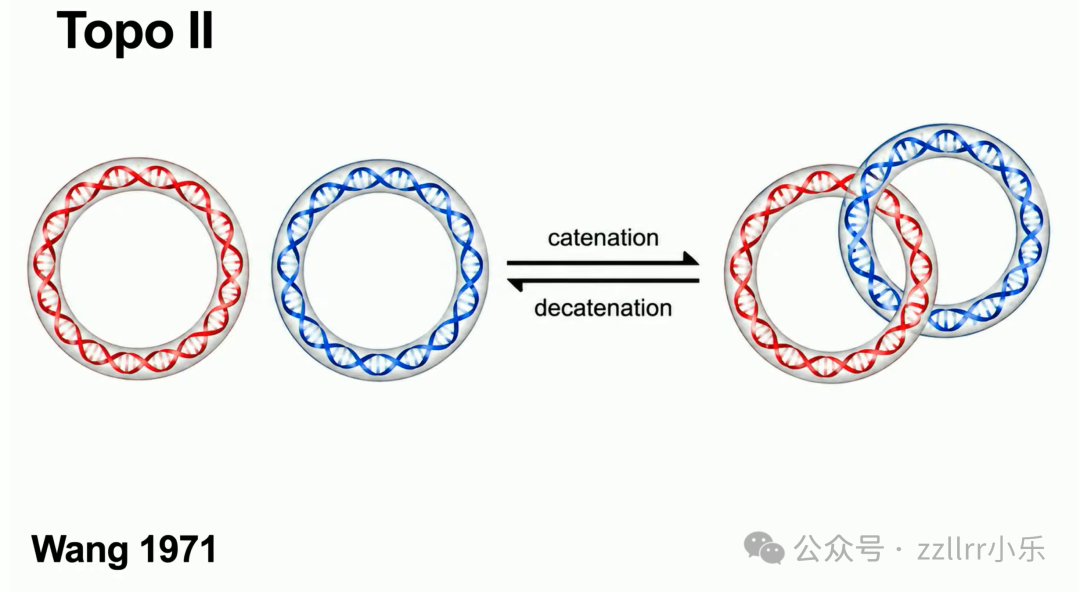
抗癌药物可抑制拓扑异构酶活性,阻断癌细胞DNA复制。
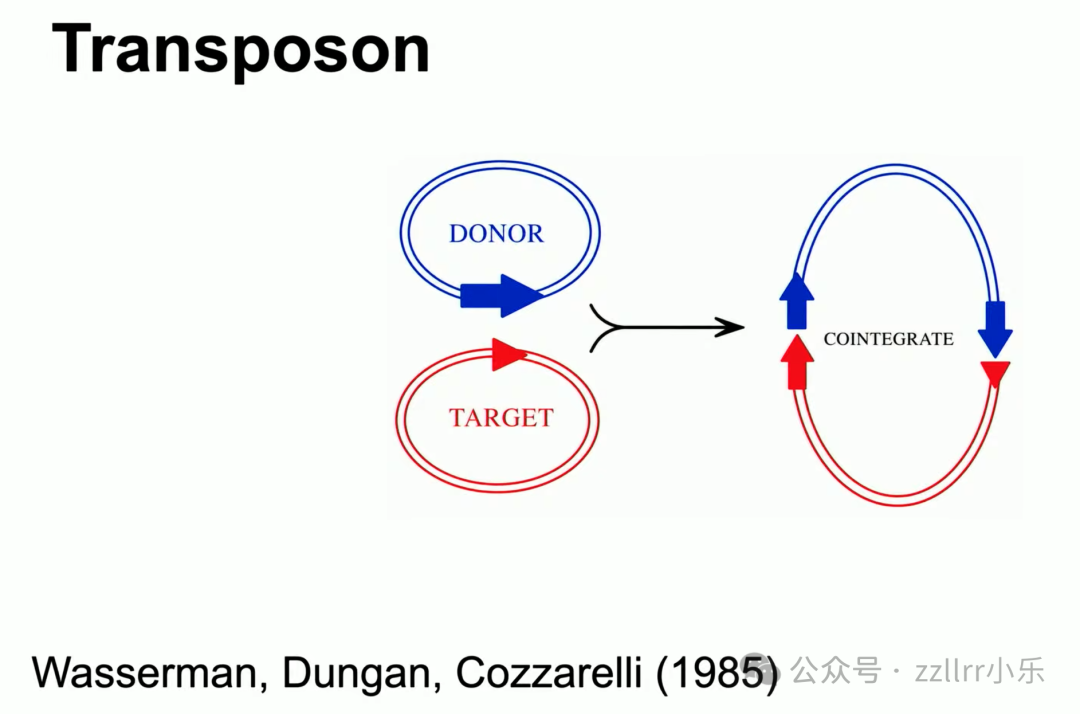
伯克利学者尼克・卡雷利(Nick Karelli)80年代利用转座子(transposon,转位子、转移子、跳跃基因)定点切割 DNA,
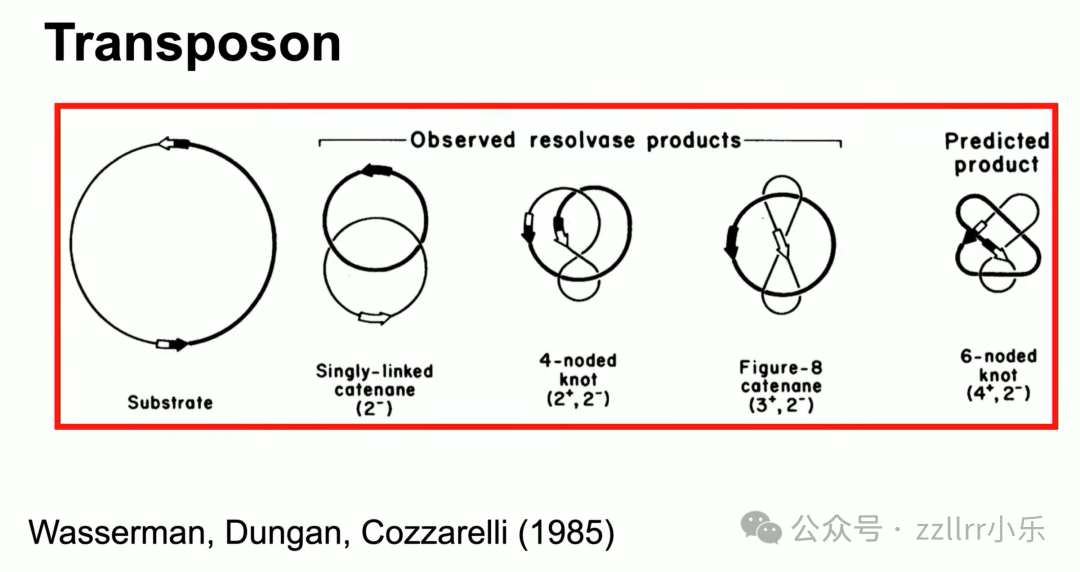

通过纽结拓扑方程预测DNA打结形态,
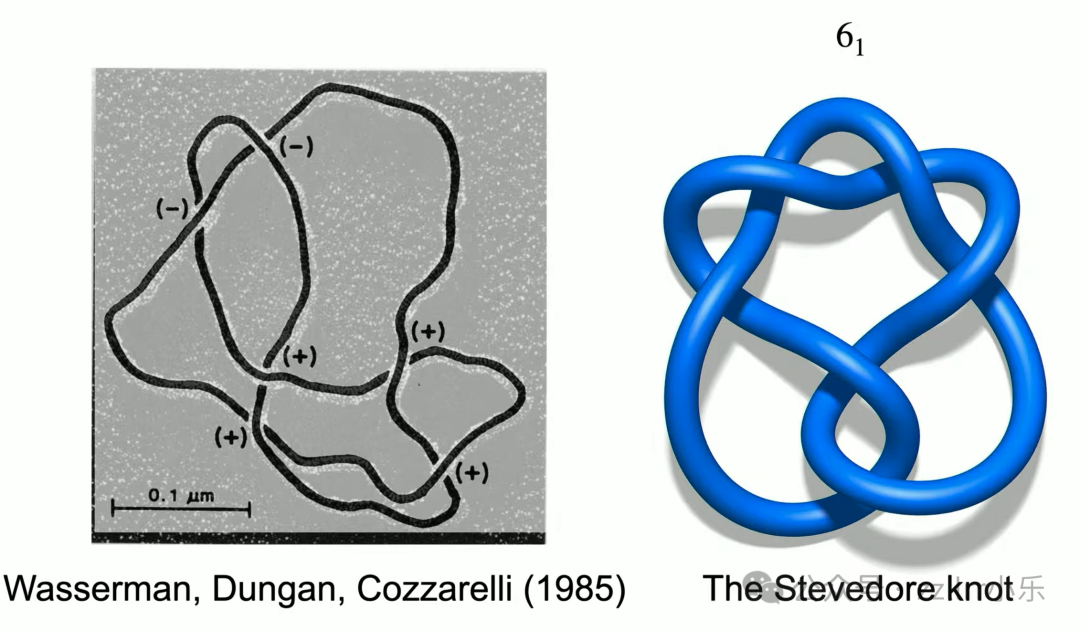
电子显微镜观测证实理论预判的6₁斯蒂夫多结(Stevedore knot),
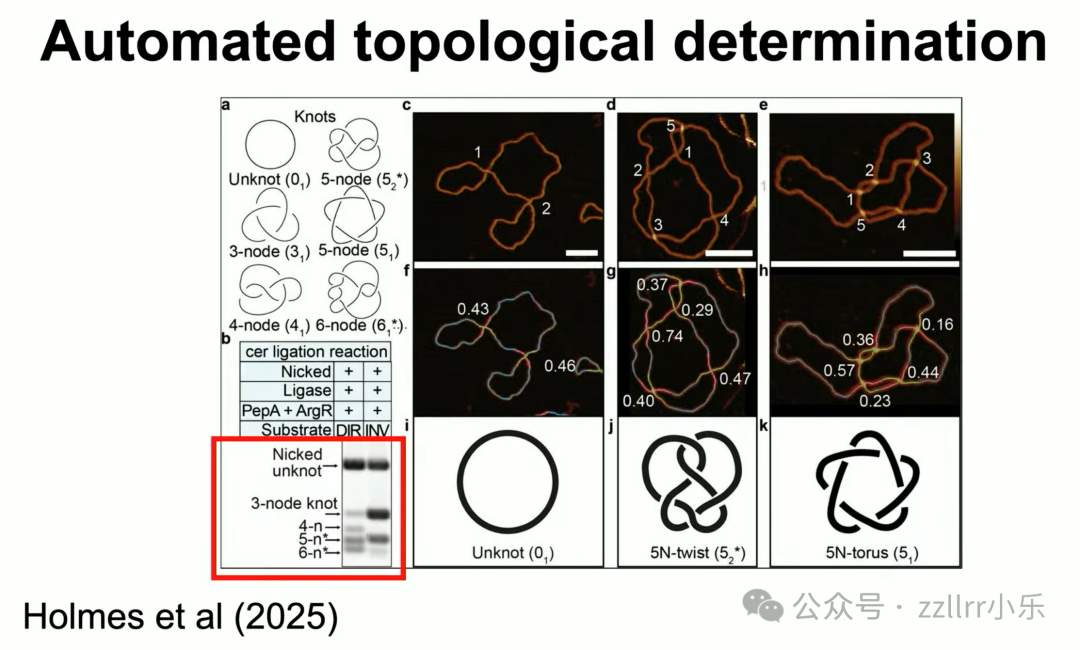
开创拓扑分子生物学全新领域。
现有原子力显微镜(atomic force microscopy)可自动识别 DNA 结类型。

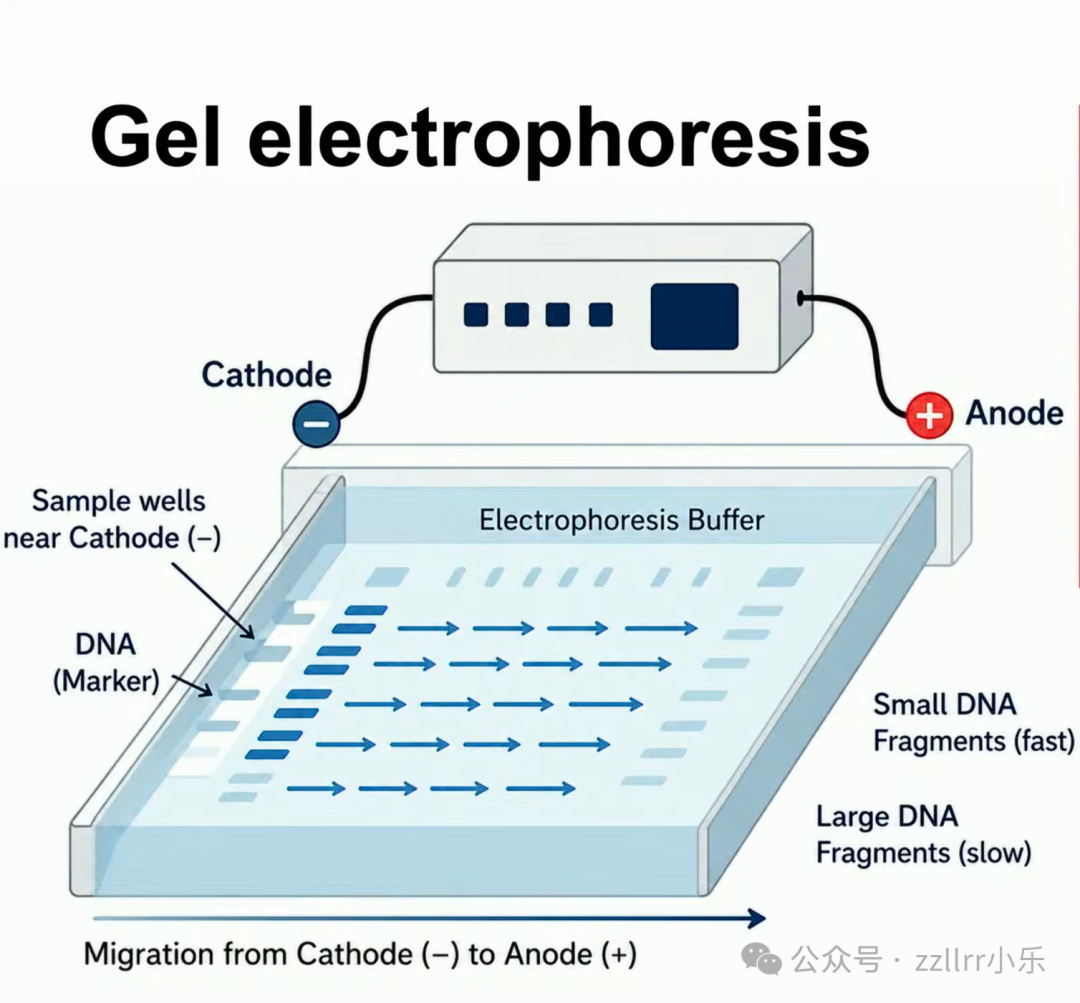
凝胶电泳(gel electrophoresis)技术可区分不同拓扑 DNA,
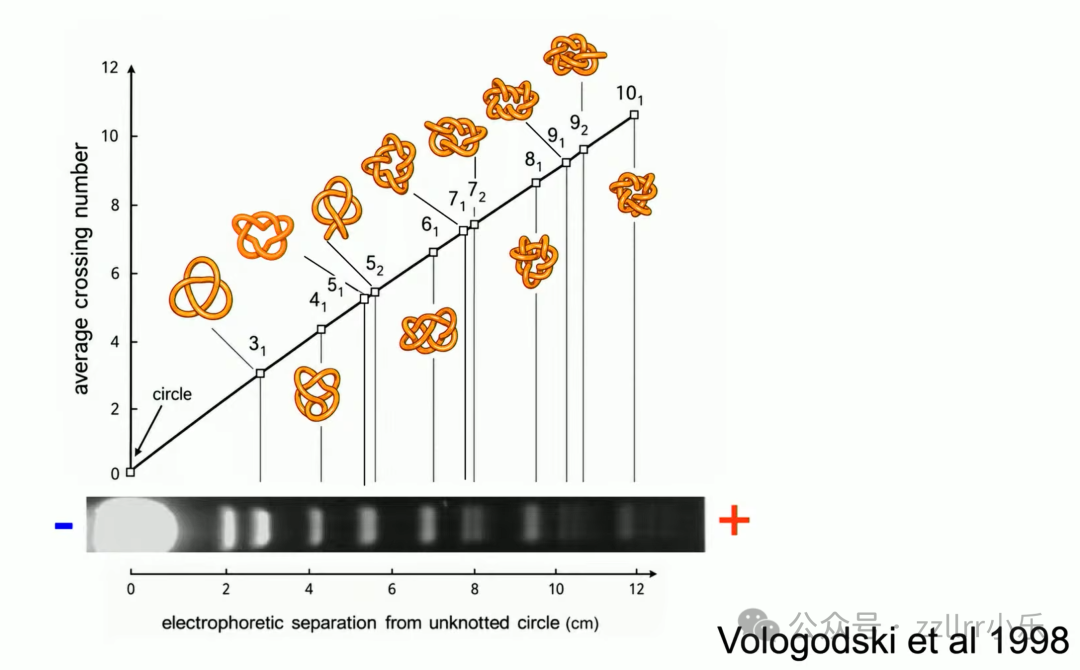
结平均交叉数越大,电泳迁移速度越慢。
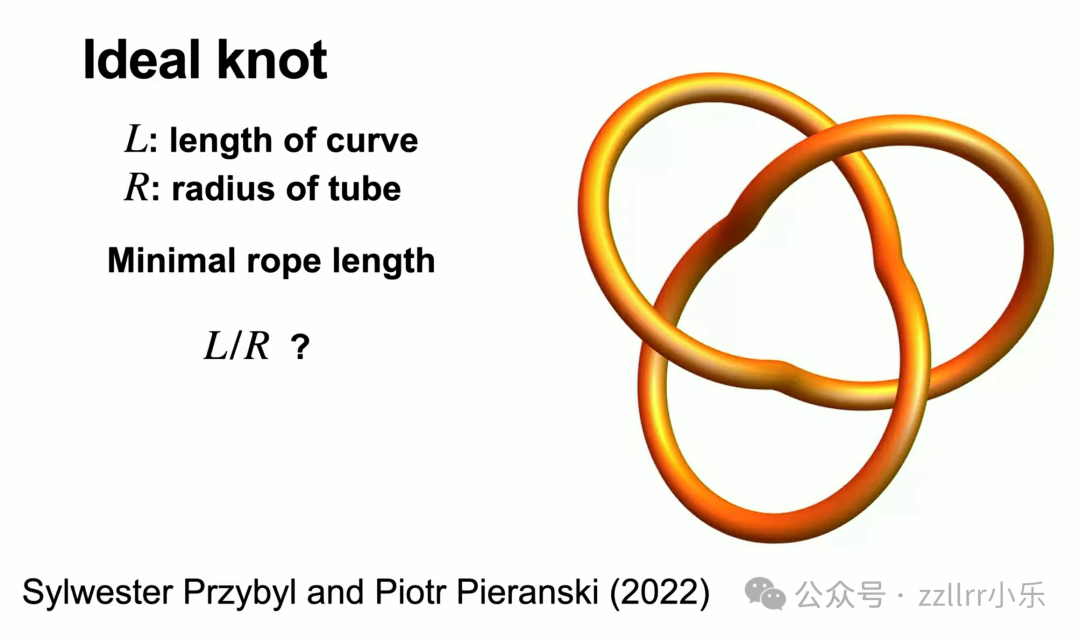
由此引出 理想结(ideal knot) 概念:

固定线材粗细,在不挤压自交前提下,能达到的最饱满构型,对应最小线材长度,例如单位半径三叶理想结最小长度为32.7429345。
瑞士联邦理工学院(EPFL)乔瓦尼・迪特勒(Giovanni Dietler)3D打印各类理想结模型,
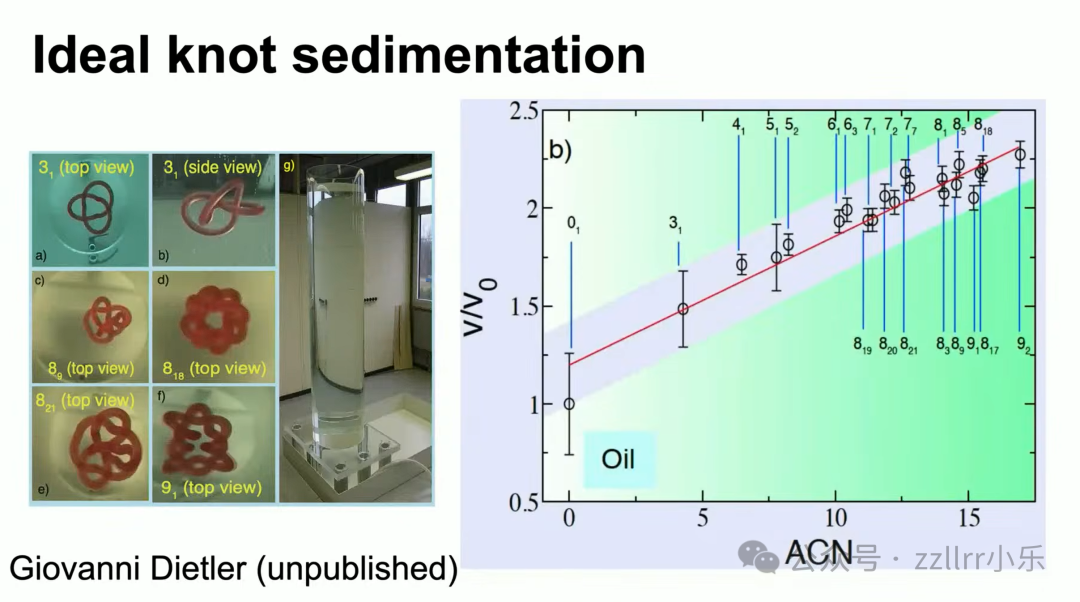
投入高粘性流体观测沉降速度,沉降速率与平均交叉数呈线性相关。
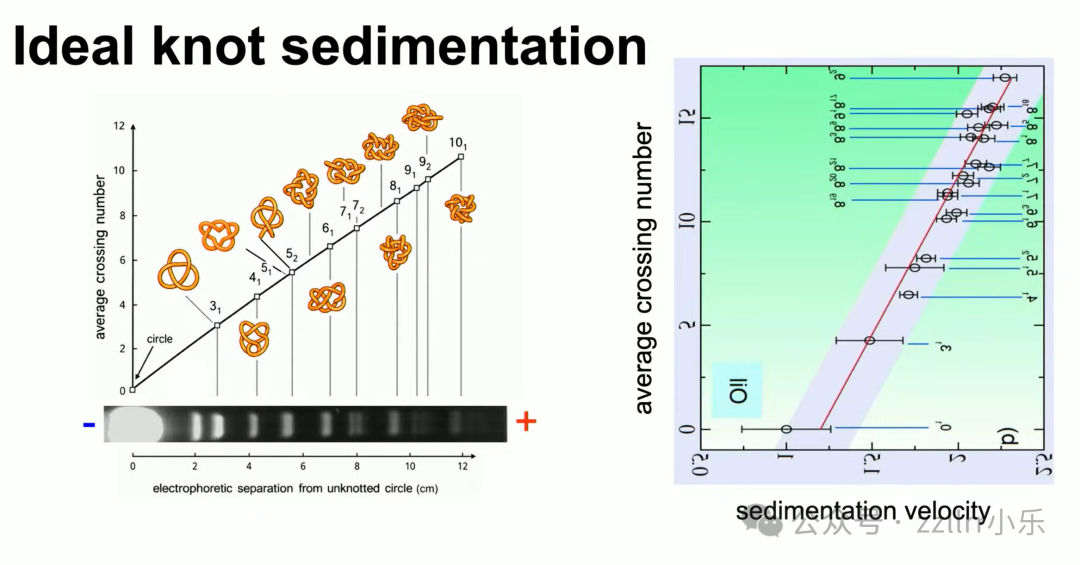
凝胶迁移、流体沉降两套物理实验,印证拓扑交叉数、几何理想结、真实DNA分子三者统一规律,环形DNA平均形态趋近理想结。
结现实应用:脐带结
24年前我第三个孩子即将出生时,我开始思考脐带打结概率。
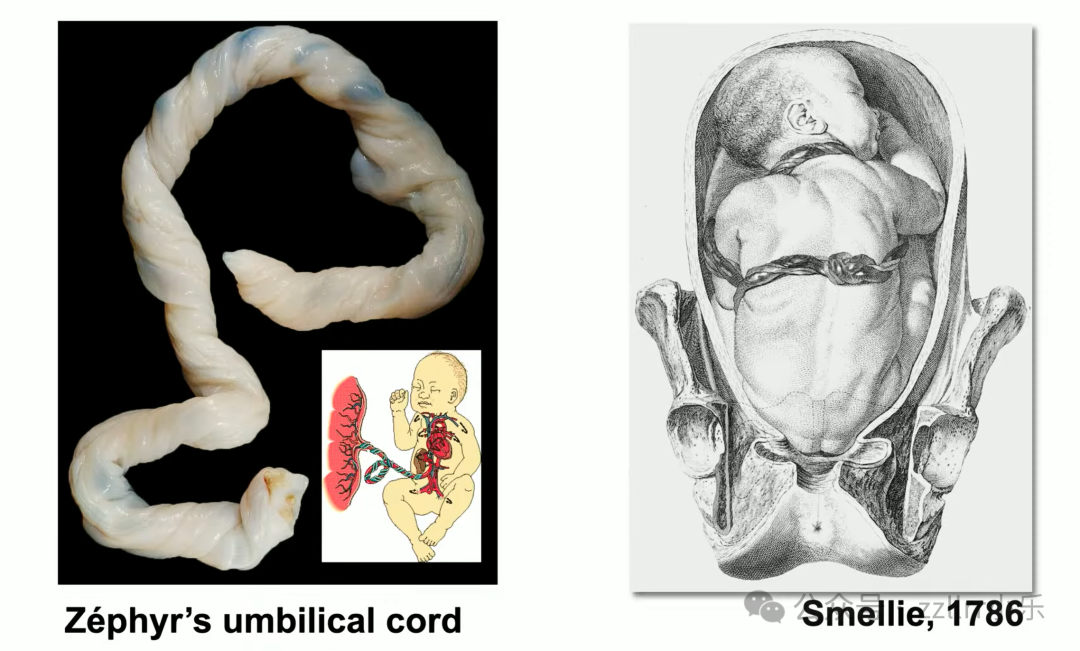
史料记载1786年威廉・斯梅利(William Smellie,1697 - 1763)便绘制脐带结案例,
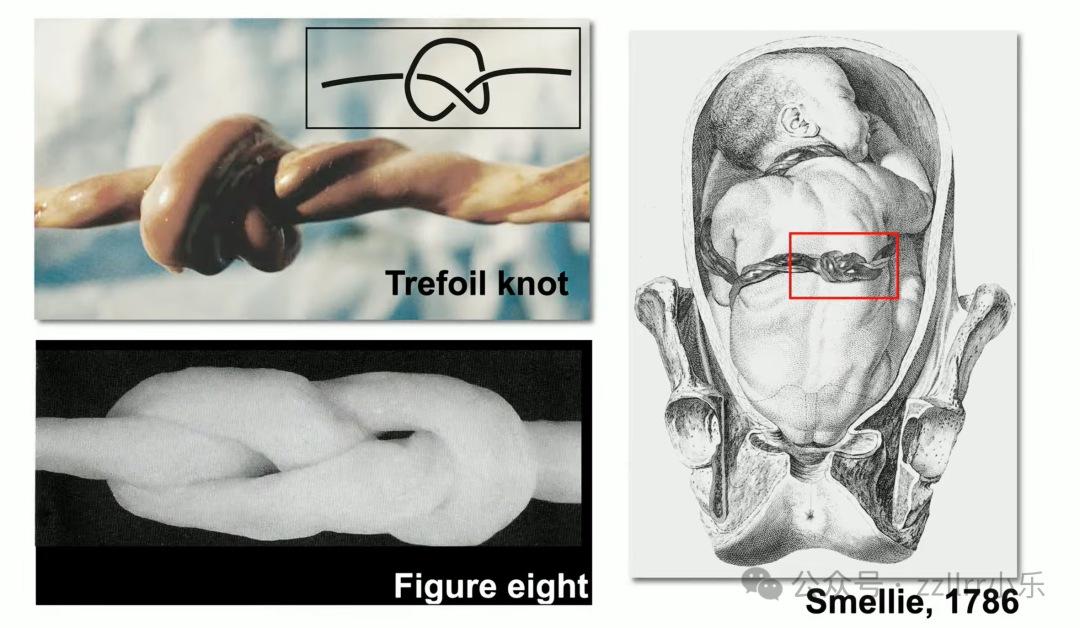
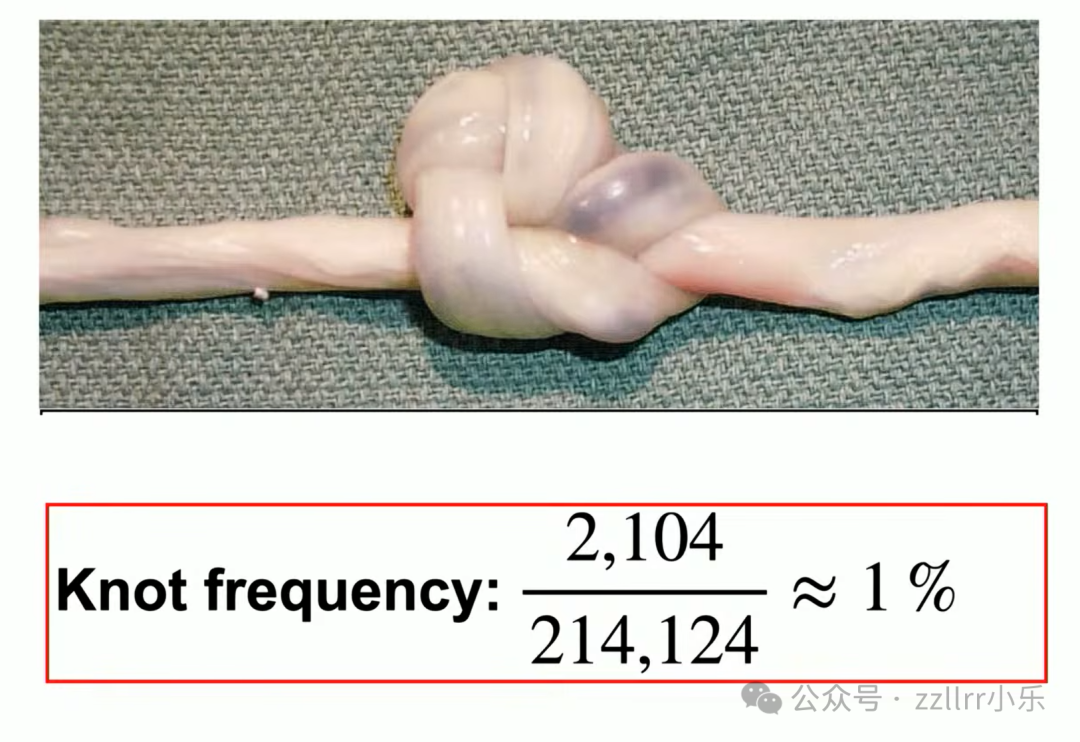
临床两万例新生儿中约两千例出现脐带真结(true knot),
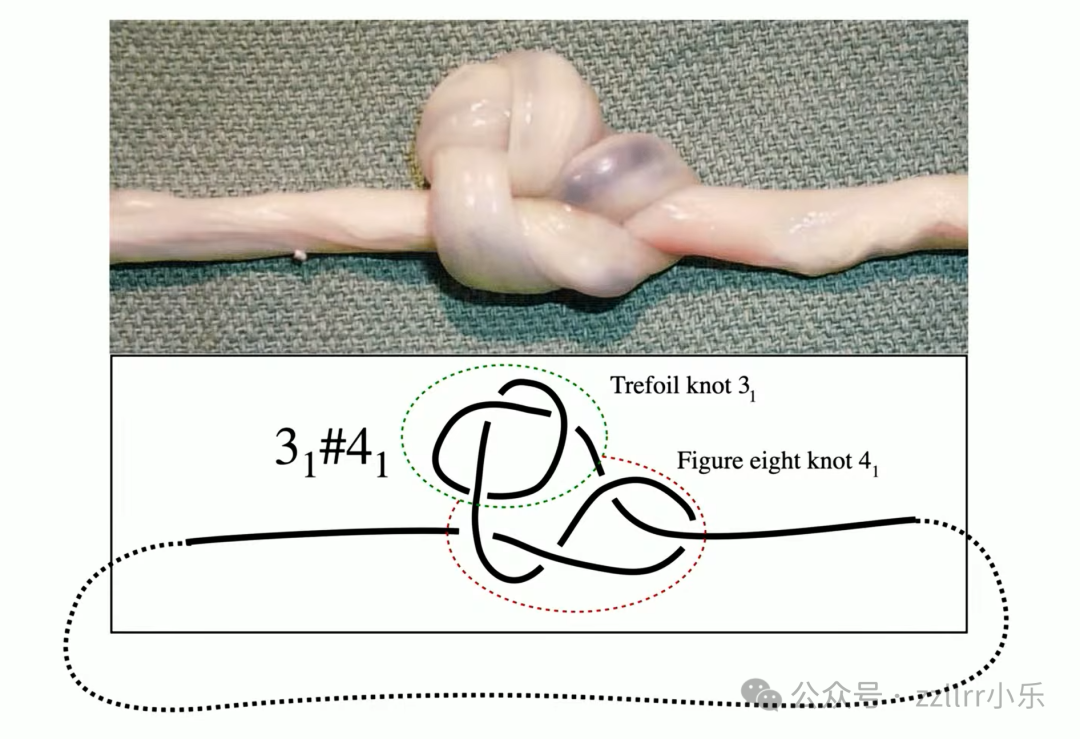
发生率1%,包含三叶结、八字结乃至复杂复合结。
模拟实验:将绳索放入密闭容器持续翻滚,线材越长打结概率越高,容器空间越小打结概率越低。
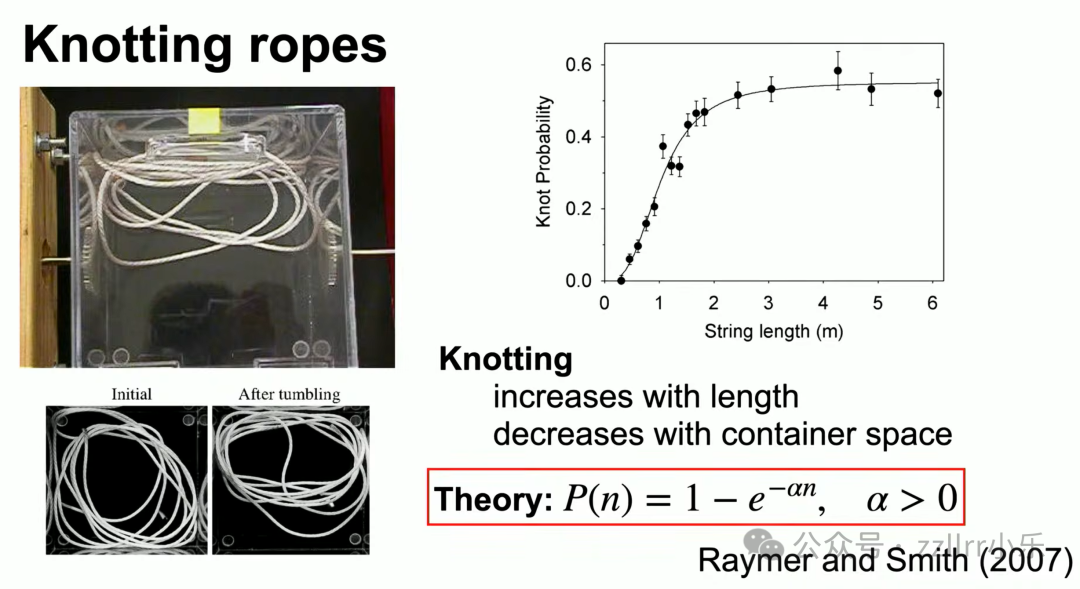
数学模型采用自回避随机游走(self-avoiding random walk),行走长度趋近无穷时打结概率无限趋近1。
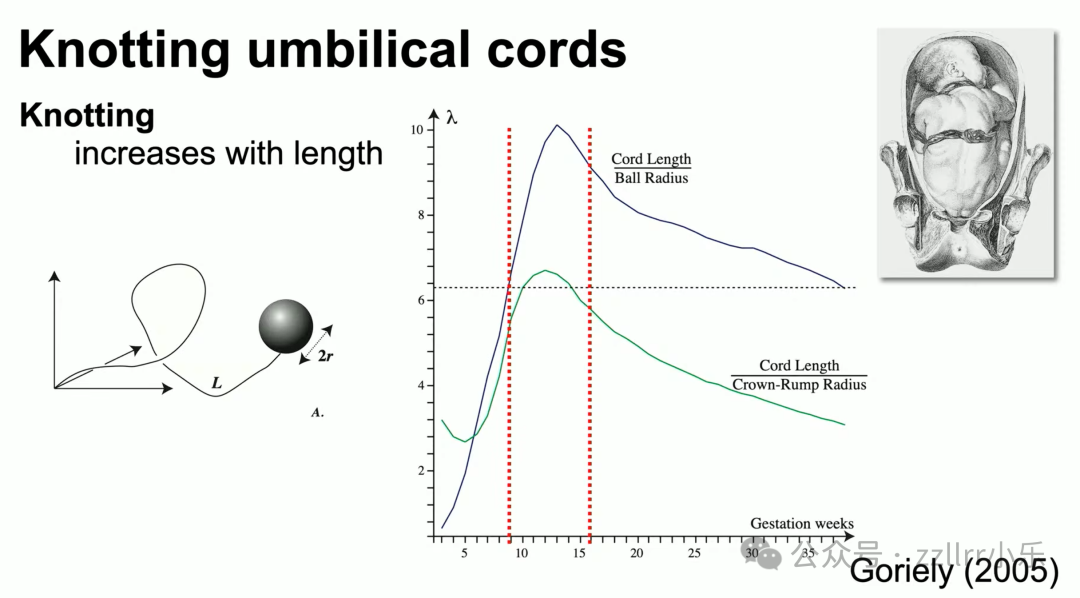
胎儿脐带打结概率关键变量:脐带长度与胎儿球体半径比值。
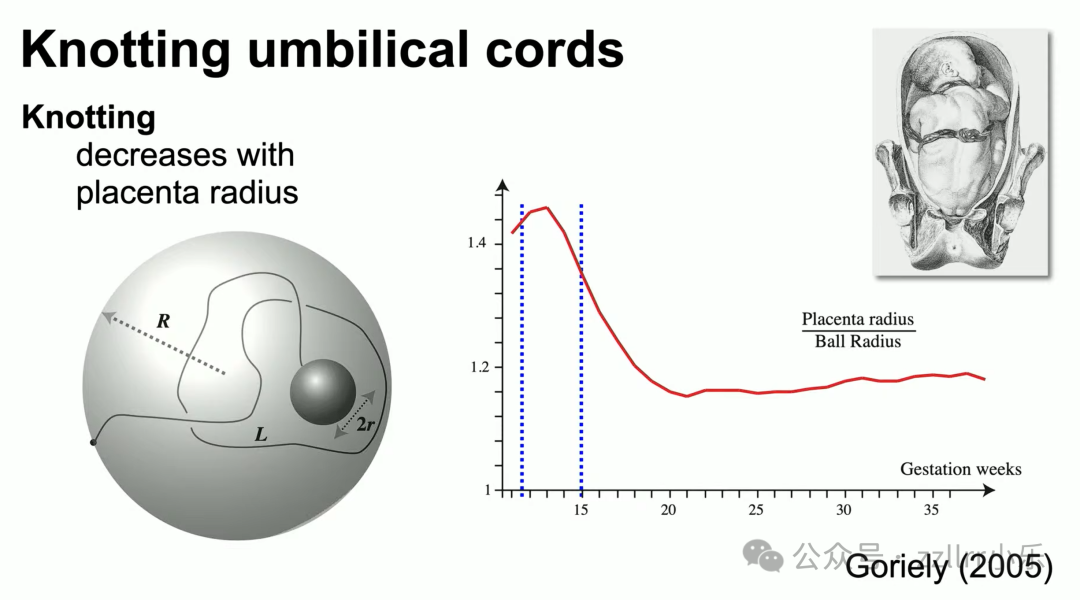
9至16周胎儿体型、脐带长度配比达到打结峰值;同卵单羊膜双胎共享胎盘羊膜,脐带易形成互锁链环,风险远高于单胎。脐带结会压迫血管阻断供血,但胎儿活跃度高往往脐带更长,整体风险可控。
结其他领域与总结

纽结理论横跨多门数学分支,已有五项菲尔兹奖相关成果,应用涵盖计算复杂度、流体涡环、天体磁通量、分子生物、植物藤蔓等。150年前彼得・泰特(Peter Tait)便预言纽结研究会衍生无穷兼具理论与实用价值的课题,欢迎大家深入探索。

Q&A 问答环节
观众问:您提到DNA近似理想结,分子生物学家能依靠该结论获得什么实际研究价值?
答:分子生物学核心课题是碱基序列如何改变DNA空间构型。理想结模型可作为标准对照,修改碱基序列后,通过模拟与凝胶观测对比形变规律,解析基因序列对分子拓扑的调控作用,为酶反应机制提供几何参考。
观众问:您所说环形DNA是细菌质粒、线粒体DNA,而非细胞核DNA吗?
答:如今可线上定制目标DNA序列,厂商直接合成纯环形样本,无需提取生物体内核酸;整套拓扑理论对质粒、线粒体、细胞核DNA全部适用,环形DNA如同分子乐高,可精准定制各类结、链环构型。
观众问:日常生活中哪些绳结缠绕需要我们留意,有无解决办法?
答:伯克利机械工程教授奥利弗・奥雷利(Oliver O'Reilly)曾发表鞋带打结论文,不同构型鞋带滑脱概率差异极大。户外徒步鞋常出现单侧易松问题,可选用高稳定性复合绳结。
观众问:授课大量实物案例,学生是否更容易理解抽象结理论?
答:因人而异,多数学生依靠现实应用建立学习兴趣;也有纯粹热爱抽象拓扑、无需具象案例的学习者,两类学习路径都合理。
观众问:纽结理论是仍存在大量重大待发现,还是学科体系已成熟?
答:该领域极为活跃,纯纽结理论有诸多悬而未决的复杂度难题;同时拓展至高维结、链环、辫结构,和多项式、曲面、积分数学深度关联,如同数论一般长期持续产出新成果。
观众问:近年基因编辑飞速发展,纽结拓扑理论对基因编辑成功率有无推动,未来会影响人类基因改造吗?
答:我仅能解读数学层面内容,伦理问题有专职教授研讨。早期拓扑模型奠定基础,如今CRISPR等新技术大幅提升基因精准调控能力。各类科技(人工智能、基因编辑)的利弊需要科学界系统推演,科研人员、普通民众、政策制定者都需建立科学认知,共同约束技术滥用。

主持人:本场系列讲座圆满收官,明年九月您将重返学院,明年课程以实操实验为主,期待更多实物演示,感谢各位到场。
参考资料
https://www.youtube.com/watch?v=UF4HU3edjwM
https://www.gresham.ac.uk/watch-now/shape-knots
https://www.gresham.ac.uk/watch-now/series/geometry-nature
本文为澎湃号作者或机构在澎湃新闻上传并发布,仅代表该作者或机构观点,不代表澎湃新闻的观点或立场,澎湃新闻仅提供信息发布平台。申请澎湃号请用电脑访问http://renzheng.thepaper.cn。



- 报料热线: 021-962866
- 报料邮箱: news@thepaper.cn
互联网新闻信息服务许可证:31120170006
增值电信业务经营许可证:沪B2-2017116
© 2014-2026 上海东方报业有限公司

